La Cour de justice de l’Union européenne tranche sur la prescription obligatoire des vaccins COVID-19 et la responsabilité des médecins
La Cour de justice de l’Union européenne (CJUE) a récemment rendu une décision qui pourrait avoir un impact majeur sur la responsabilité des professionnels de santé ayant administré les vaccins contre le COVID-19. Selon cet arrêt, une prescription médicale était nécessaire pour l’administration de ces vaccins, ce qui remet en cause la manière dont la campagne de vaccination a été menée dans plusieurs pays européens.
L’importance de la prescription médicale
L’affaire portée devant la CJUE par le professeur Frajese met en lumière un point fondamental : les médecins avaient la liberté d’évaluer la pertinence d’une vaccination pour leurs patients. Contrairement à ce qui a été appliqué dans certains États membres, l’autorisation de mise sur le marché des vaccins COVID-19 n’impliquait aucune obligation pour les médecins de prescrire ou d’administrer ces vaccins.
La Cour rappelle que la relation médecin-patient repose sur un principe essentiel : la liberté de prescription. Chaque professionnel de santé doit pouvoir, en fonction des spécificités du patient, choisir le traitement le plus adapté, en toute indépendance. Ainsi, ceux qui ont choisi de ne pas recommander ces vaccins ou de les déconseiller ne peuvent être poursuivis ni disciplinés pour cette raison.
Les conséquences sur la responsabilité des médecins
L’arrêt de la CJUE précise que les médecins ayant administré ces vaccins sans évaluation individuelle du patient pourraient voir leur responsabilité engagée. En effet, le fait que ces vaccins aient reçu une autorisation de mise sur le marché (AMM) ne suffisait pas à exempter les praticiens de leur devoir d’analyse des risques pour chaque patient.
Cela signifie que les professionnels de santé qui ont procédé à la vaccination sans prescription spécifique pourraient être poursuivis pour faute médicale. La Cour souligne que la protection juridique dont bénéficiaient les médecins ne s’applique que si l’administration des vaccins a respecté les règles en vigueur, ce qui inclut l’évaluation individuelle de chaque patient.
L’impact sur les recours en justice
Les décisions de la CJUE s’imposent aux juges nationaux, et cet arrêt pourrait influencer de nombreuses procédures en cours. En particulier, les personnels de santé suspendus ou radiés pour avoir refusé d’administrer ces vaccins pourraient voir leur situation réexaminée à la lumière de cette jurisprudence.
Par ailleurs, les personnes ayant subi des effets indésirables après la vaccination disposent désormais d’un argument supplémentaire pour engager des actions en responsabilité contre les médecins et les autorités sanitaires. L’absence de prescription médicale constitue une violation des règles établies, rendant possibles des demandes d’indemnisation pour préjudice corporel.
Une décision qui redéfinit la liberté de soins
Cette décision de la CJUE marque un tournant majeur dans l’approche des politiques de santé publique. Elle réaffirme le principe selon lequel les médecins doivent conserver leur liberté de prescription et rappelle que toute obligation de vaccination doit être encadrée par des règles claires respectant les droits des patients.
Alors que les débats sur les politiques vaccinales continuent, cette décision apporte une clarification essentielle : les professionnels de santé doivent être libres de prescrire ou de déconseiller un traitement, en fonction des besoins réels de leurs patients, sans subir de pressions extérieures.
Si vous êtes victime d’un effet secondaire suite à la vaccination, il est important de savoir que vous avez des droits.Lecabinet ELLIPSIS AVOCATS se spécialise dans la défense des victimes de dommages corporels et vous accompagne pour obtenir une indemnisation et faire valoir vos droits face aux autorités compétentes.
N’hésitez pas à nous contacter pour toute question ou pour discuter de votre situation.
Certainement un des ouvrages les plus complets et sourcés sur « l’affaire Covid ». Une enquête d’utilité publique qui se lit comme un polar. Un livre à faire circuler de toute urgence.
Un vague sentiment de vous être fait avoir ? Pourquoi ? Comment ? Fruit d’une enquête minutieuse, ce livre choc vous dévoile les coulisses et les mensonges des « pandémies », présentes, passées et à venir. Des faits, rien que des faits. Qui a orchestré cette fraude planétaire, quand et comment les membres d’un petit groupe organisé ont patiemment tissé leur toile, corrompu le système et subverti les représentants des peuples ? Dix grands mensonges, dix grandes vérités. Voici enfin la vérité, voici les réponses.
Soyez prêts à protéger votre santé, et, peut-être, sauver votre vie.
Le projet mondial sur la sécurité des vaccins contre la COVID-19 (GCoVS), lancé en 2021 dans le cadre du Réseau mondial de données sur les vaccins (GVDN®), vise à évaluer l’innocuité des vaccins. Cette étude visait à évaluer le risque d’ effets indésirables particuliers (EIPP) suite à la vaccination contre la COVID-19 sur dix sites répartis dans huit pays.
Méthodes
À l’aide d’un protocole commun, cette étude de cohorte observationnelle a comparé les taux observés et attendus de 13 effets indésirables graves (EI) sélectionnés pour les résultats neurologiques, hématologiques et cardiaques. Les taux attendus ont été obtenus par les sites participants à partir des données de santé pré-vaccinales COVID-19, stratifiées par âge et par sexe. Les taux observés ont été rapportés à partir des mêmes bases de données de santé depuis le déploiement du programme de vaccination contre la COVID-19. Les EI survenant jusqu’à 42 jours après la vaccination par les vaccins à ARNm (BNT162b2 et ARNm-1273) et à vecteur adénoviral (ChAdOx1) ont été inclus dans l’analyse principale. Les risques ont été évalués à l’aide des ratios observés/attendus (OE) avec des intervalles de confiance à 95 %. Les signaux de sécurité potentiels prioritaires étaient ceux dont la limite inférieure de l’intervalle de confiance à 95 % (LBCI) était supérieure à 1,5.
Résultats
Français Les participants comprenaient 99 068 901 personnes vaccinées. Au total, 183 559 462 doses de BNT162b2, 36 178 442 doses d’ARNm-1273 et 23 093 399 doses de ChAdOx1 ont été administrées dans les sites participants au cours de la période d’étude. Les périodes à risque suivant les calendriers de vaccination homologues ont contribué à 23 168 335 années-personnes de suivi. Des ratios OE avec LBCI > 1,5 ont été observés pour le syndrome de Guillain-Barré (2,49, IC à 95 % : 2,15, 2,87) et la thrombose du sinus veineux cérébral (3,23, IC à 95 % : 2,51, 4,09) après la première dose du vaccin ChAdOx1. L’encéphalomyélite aiguë disséminée a montré un ratio d’efficacité de 3,78 (IC à 95 % : 1,52, 7,78) après la première dose du vaccin à ARNm-1273. Les ratios d’efficacité de la myocardite et de la péricardite après BNT162b2, ARNm-1273 et ChAdOx1 ont augmenté significativement avec des LBCI > 1,5.
Conclusion
Cette analyse multi-pays a confirmé les signaux de sécurité préétablis concernant la myocardite , la péricardite , le syndrome de Guillain-Barré et la thrombose du sinus veineux cérébral . D’autres signaux de sécurité potentiels nécessitant une investigation plus approfondie ont été identifiés.
Questions répondues dans cet article
Bêta propulsé par GenAI
Il s’agit d’un contenu d’IA génératif et la qualité peut varier. Apprendre encore plus .
Événements indésirables présentant un intérêt particulier
COVID 19
Analyse observée vs. attendue
1. Introduction
Depuis la déclaration de la pandémie de COVID-19 par l’Organisation mondiale de la santé (OMS) le 11 mars 2020 [1], plus de 13,5 milliards de doses de vaccins contre la COVID-19 ont été administrées dans le monde [2] . En novembre 2023, au moins 70,5 % de la population mondiale avait reçu au moins une dose d’un vaccin contre la COVID-19 [2] . Ce scénario sans précédent souligne le besoin urgent d’une surveillance complète de la sécurité des vaccins, car de très rares événements indésirables associés aux vaccins contre la COVID-19 pourraient n’être révélés qu’après l’administration à des millions de personnes.
Français En prévision de ce déploiement mondial sans précédent de vaccins contre la COVID-19, l’initiative Safety Platform for Emergency vACcines (SPEAC) a formulé une liste d’événements indésirables potentiels du vaccin contre la COVID-19 présentant un intérêt particulier (AESI) en 2020 [3] . La sélection des AESI était basée sur leurs associations préétablies avec la vaccination, des plateformes vaccinales ou des adjuvants spécifiques, ou la réplication virale au cours d’une maladie de type sauvage ; des préoccupations théoriques liées à l’immunopathogenèse ; ou des preuves à l’appui provenant de modèles animaux utilisant des plateformes vaccinales candidates [3] .
Français Une approche flexible pour évaluer les EISP est la comparaison des taux d’EISP observés après l’introduction d’un programme de vaccination avec les taux attendus (ou de fond) basés sur des périodes historiques avant le déploiement du vaccin [4] , [5] . De telles comparaisons peuvent être effectuées rapidement et peuvent jouer un rôle clé dans la détection précoce de signaux potentiels de sécurité des vaccins ou lorsque les agences de réglementation et de santé publique ont besoin d’une évaluation rapide d’un signal de sécurité émergent [4] , [6] . L’analyse observée versus (vs.) attendue (OE) a joué un rôle essentiel dans l’identification de la thrombose avec syndrome de thrombocytopénie (TTS) comme un signal de sécurité, ce qui a conduit à la suspension de l’utilisation du ChAdOx1 (vaccin AstraZeneca COVID-19) le 11 mars 2021, au Danemark et en Norvège [7] , [8] .
Français Ces évaluations sont utiles non seulement au début du déploiement à grande échelle des vaccins, mais aussi à mesure que le programme de vaccination mûrit, en particulier si elles peuvent être menées dans un contexte multinational. Nous avons mené une étude de cohorte mondiale en suivant le protocole d’étude Observed vs. Expected Analyses of COVID-19 Adverse Events of Special Interest [9] avec des données provenant de 10 sites dans huit pays participant au projet unique Global COVID Vaccine Safety (GCoVS) [10] du Global Vaccine Data Network™ (GVDN®) [11] . Le projet GCoVS, lancé en 2021, est une collaboration mondiale financée par les Centers for Disease Control and Prevention (CDC) entre des chercheurs et des sources de données de plusieurs pays à des fins de surveillance de la sécurité des vaccins contre la COVID-19.
2. Méthodes
2.1 . Conception de l’étude
Cette étude observationnelle rétrospective a été conçue pour estimer les ratios OE d’EIS sélectionnés après la vaccination contre la COVID-19 dans une cohorte de population multi-pays.
2.2 . Source des données et population étudiée
Le projet GCoVS a compilé des données de santé électroniques sur les effets indésirables graves (EIS) liés aux vaccins contre la COVID-19 auprès de participants de plusieurs sites du réseau GVDN, notamment en Argentine, en Australie (Nouvelle-Galles du Sud), en Australie (Victoria), au Canada (Colombie-Britannique), au Canada (Ontario), au Danemark, en Finlande, en France, en Nouvelle-Zélande et en Écosse [10] . Les données de santé comprenaient des données individuelles ou populationnelles, selon la disponibilité dans les sites d’étude ( tableau supplémentaire 1 ).
La majorité des sites d’étude ont utilisé des registres de vaccination contenant des données individuelles. Ces registres couvraient la même population et la même région géographique que les ensembles de données utilisés pour calculer les taux de base. Nous avons également examiné les données populationnelles sur la couverture vaccinale à l’aide de tableaux de bord régulièrement mis à jour par les sites d’étude. Si le nombre de personnes vaccinées par tranches d’âge et de sexe spécifiques était disponible, nous avons converti ces chiffres en années-personnes en fonction de la période de risque post-vaccinale. Contrairement aux registres contenant des données individuelles, les strates d’âge et de sexe utilisées dans cette approche pourraient ne pas correspondre à celles utilisées pour le calcul des taux de base.
Les participants étaient des personnes vaccinées contre la COVID-19 au sein des populations représentées par les sites. Dans la mesure du possible, des méthodes standardisées ont été appliquées entre les sites. Les types de patients comprenaient des patients hospitalisés (Australie – Nouvelle-Galles du Sud, France, Nouvelle-Zélande, Écosse) et des patients hospitalisés et ambulatoires aux urgences (Argentine, Australie – Victoria, Canada, Danemark, Finlande). Dans les pays où les types de patients n’étaient pas clairement définis, la durée du contact à l’hôpital a été utilisée comme indicateur indirect des types de patients. À titre d’exemple, une durée de contact de cinq heures ou plus a été utilisée comme indicateur indirect des patients hospitalisés au Danemark. Les caractéristiques spécifiques des sources de données et des données par site sont présentées dans le tableau supplémentaire 1 .
2.3 . Période d’étude et suivi
Français Les périodes d’étude variaient selon les pays, commençant à la date du déploiement du programme de vaccination contre la COVID-19 spécifique au site et se terminant à la fin de la disponibilité des données ( tableau 1 ). En général, les périodes d’étude s’étendaient de décembre 2020 à août 2023. La période d’étude la plus courte observée a eu lieu en Australie – Nouvelle-Galles du Sud, comprenant 11 mois de février 2021 à décembre 2021. L’Argentine a eu la période d’étude la plus longue, de décembre 2020 à août 2023, comprenant un total de 32 mois.
Tableau 1. Résumé de la population par site. (Uniquement les vaccins Pfizer/BioNTech BNT162b2, Moderna mRNA-1273 et Oxford/Astra Zeneca/Serum Institute of India ChAdOx1 et les doses 1 à 4 inclus).
Vaccins : Pfizer/BioNTech (BNT162b2), Moderna (ARNm-1273) et Oxford/Astra Zeneca/Serum Institute of India (ChAdOx1).
Français Les intervalles de risque utilisés après chaque dose étaient de 0 à 7 jours, de 8 à 21 jours, de 22 à 42 jours et de 0 à 42 jours. Pour chaque dose de vaccination, le jour 0 était le jour de la réception du vaccin. Pour ce manuscrit, nous présentons les résultats pour l’intervalle de risque de 0 à 42 jours uniquement. D’autres données sont présentées sur le tableau de bord du GVDN avec toutes les dernières mises à jour des sites participants [12] . Les événements survenus en dehors de la période d’étude n’ont pas été inclus. Une période de sevrage de 365 jours pour les événements survenus a été utilisée pour définir les événements incidents. Les événements survenus étaient considérés comme incidents s’il n’y avait pas d’enregistrement du même événement survenu au cours de la période de sevrage de 365 jours précédente. Un individu peut avoir contribué à plusieurs événements survenus à condition qu’ils soient séparés dans le temps par au moins la période de sevrage de 365 jours.
2.4 . Variables et résultats de l’étude
2.4.1 . Événements indésirables présentant un intérêt particulier (EIIP)
En novembre 2023, plusieurs vaccins contre la COVID-19 étaient utilisés par les sites GCoVS, représentant plusieurs types de plateformes, telles que les plateformes à vecteurs viraux inactivés, à base d’acide nucléique (ARNm), à base de protéines et non réplicatifs ( tableau 2 ). Pour ce manuscrit, nous nous sommes concentrés sur les trois vaccins ayant enregistré le plus grand nombre de doses administrées : Pfizer/BioNTech BNT162b2, Moderna mRNA-1273 et Oxford/Astra Zeneca/Serum Institute of India ChAdOx1. Français Le nombre cumulé de doses d’autres vaccins administrées (n) dans les sites d’étude était relativement faible, à l’exception des vaccins inactivés Sinopharm (n = 134 550) et Sinovac (n = 31 598), du vaccin à base de protéines Novavax (n = 66 856) et des vaccins à vecteur adénoviral Janssen/Johnson & Johnson (n = 1 137 505) et Gamaleya Research Institute/Sputnik (n = 84 460). Le nombre total de doses de chaque marque de vaccin administrées est indiqué dans le tableau 2. Les données sur l’exposition au vaccin contre la COVID-19 par plateforme/type, marque et dose étaient disponibles au niveau individuel pour déterminer le nombre de cas observés par type/marque de vaccin et profil de dose et dans l’intervalle de risque post-vaccination de 0 à 42 jours.
Tableau 2. Nombre total de vaccinations par marque.
Plateforme de vaccins
Marque de vaccin
Doses totales
Inactivé
Vaccin Covilo ou SARS-CoV-2 (cellule Vero) [Sinopharm (Pékin)]
134 550
Covaxin [Bharat Biotech]
1 660
CoronaVac ou Sinovac [Sinovac Biotech]
31 598
Inactivé (cellule Vero) [Sinopharm (Wuhan)]
623
À base d’acide nucléique
Vaccin bivalent Comirnaty ou Riltozinaméran ou Pfizer/BioNTech contre la COVID-19 [Pfizer/BioNTech]
3 516 963
Comirnaty ou Tozinameran [Pfizer/BioNTech ou Fosun-BioNTech]
183 677 660
Comirnaty ou Tozinameran pédiatrique [Pfizer/BioNTech ou Fosun-BioNTech]
2 439 086
Spikevax bivalent Original/Omicron [Moderna]
2 750 476
Demi-dose d’élasomeran ou de Spikevax ou de TAK-919 [Moderna ou Takeda]
400 395
Élasomérane ou Spikevax ou TAK-919 [Moderna ou Takeda]
36 222 514
À base de protéines
MVC-COV1901 [Medigen]
16
Covovax ou Nuvaxoid [Novavax ou Serum Institute of India]
66 856
Vecteur viral non réplicatif
Convidacia ou Convidance [CanSino]
3 938
Covishield ou Vaxzevria [AstraZeneca ou Serum Institute of India]
23 094 620
Spoutnik Light ou Gam-COVID-Vac [Institut de recherche Gamaleya]
26
Spoutnik V [Institut de recherche Gamaleya]
84 460
Janssen [Janssen/Johnson & Johnson]
1 137 505
2.5 . Analyse statistique
2.5.1 . Calcul des ratios observés/attendus pour chaque site
Français Pour chaque site, nous avons calculé le nombre observé d’événements pour chaque EISP dans l’intervalle de risque après l’introduction de la vaccination contre la COVID-19. Pour calculer le nombre attendu de cas, nous avons utilisé les données sur les taux de base pré-vaccination contre la COVID-19 de 2015 à 2019 (2019-2020 pour le Danemark) collectées dans l’étude GCoVS Background Rates of EISP Following COVID-19 vaccination [13] . La période de suivi observée en personnes-années pour un profil de vaccination donné et la période post-vaccinale a été stratifiée selon le groupe d’âge et le sexe. Chacune des personnes-années stratifiées selon l’âge et le sexe a été multipliée par le taux de base stratifié selon l’âge et le sexe correspondant. Cela a donné le nombre attendu de cas dans chaque strate, qui ont ensuite été additionnés pour donner le nombre total de cas attendus pendant la période de suivi observée.
Les ratios d’efficacité globale agrégés par dernière dose ont été calculés en divisant le nombre de cas observés par le nombre de cas attendus après la vaccination ; des intervalles de confiance (IC) à 95 % ont été obtenus selon la distribution exacte de Poisson. Nous avons également calculé les ratios d’efficacité globale pour les schémas homologues des vaccins BNT162b2, mRNA-1273 et ChAdOx1 jusqu’à quatre doses. Les ratios d’efficacité globale agrégés et spécifiques aux schémas homologues sont présentés.
We considered an OE ratio a potential safety signal of concern where the lower bound of the 95 % CI (LBCI) was greater than one and reached statistical significance [5]. However, we prioritised potential safety signals of concern for further evaluation where the LBCI was greater than 1.5, due to increased statistical evidence and the higher likelihood of being a true signal, based on expert opinion from the CDC and GVDN collaborators.
2.5.2 . Combinaison des résultats entre sites
Les résultats ont été agrégés sur l’ensemble des sites en additionnant le nombre d’événements observés pour chaque EISP et les années-personnes stratifiées selon l’âge et le sexe pour un profil de vaccination et une période post-vaccinale donnés. Pour chaque EISP, les profils vaccinaux individuels ont été rapportés si le nombre cumulé de suivis (en années-personnes) dans la période de 0 à 42 jours après la vaccination était de 10 000 ou plus. Le nombre combiné d’événements et le ratio d’événements indésirables ont été calculés avec des IC à 95 % dérivés de la distribution exacte de Poisson. Aucun événement (c’est-à-dire zéro) observé pour une marque de vaccin et un profil de dose n’a été rapporté séparément sans IC.
2.5.3 . Analyse de sensibilité
Premièrement, nous avons réalisé des analyses de sensibilité spécifiques à chaque site afin d’explorer plus en détail les associations potentielles entre les signaux de sécurité les plus significatifs identifiés dans l’analyse principale. Les taux observés rapportés par site ont été pris en compte dans l’analyse en fonction des contraintes suivantes. Pour chaque marque de vaccin, profil de dose et combinaison de période post-vaccinale, les ratios d’événements indésirables graves et l’IC à 95 % ont été supprimés si moins de cinq événements étaient observés. Deuxièmement, nous avons réalisé une analyse complémentaire incluant d’autres vaccins et doses administrés entre les sites. Le seuil de déclaration en personnes-années a été abaissé de 10 000 à 1 000 personnes-années par rapport à l’analyse principale des ratios d’événements indésirables graves agrégés, ce qui a permis d’analyser un plus large éventail de vaccins.
2.6 . Approbation éthique
L’approbation des comités d’éthique de la recherche humaine compétents a été obtenue ou une exemption a été obtenue pour tous les sites participants ( tableau supplémentaire 3 ).
3. Résultats
Français La population totale vaccinée sur l’ensemble des sites comprenait 99 068 901 personnes. La plupart des vaccinés appartenaient aux tranches d’âge de 20 à 39 ans et de 40 à 59 ans ( Tableau 1 ). Au total, 183 559 462 doses de BNT162b2, 36 178 442 doses d’ARNm-1273 et 23 093 399 doses de ChAdOx1 ont été administrées sur l’ensemble des sites au cours des périodes d’étude. Le plus grand nombre de doses a été administré en France (120 758 419), suivie du Canada – Ontario (32 159 817) et de l’Australie – Victoria (15 617 627). Au total, 23 168 335 personnes-années ont contribué aux ratios OE pour l’AESI suivant les calendriers homologues. Le résumé de la population est présenté dans le tableau 1 , et des informations plus détaillées sur les autres vaccins administrés sont présentées dans le tableau supplémentaire 4. Dans les sections de résultats ci-dessous, nous fournissons à la fois les ratios d’efficacité globale agrégés ( tableau 3 , tableau 4 , tableau 5 ) et les ratios d’efficacité globale détaillés pour les calendriers homologues ( figure 1 , figure 2 , figure 3 ), y compris le nombre d’événements et de personnes-années. Globalement, 95,8 % et 86,6 % des vaccinations ont été incluses dans l’analyse des calendriers agrégés et homologues, respectivement ( Tableau supplémentaire 5 ). Les principaux résultats des sites individuels ainsi que les périodes de risque supplémentaires et les méta-analyses pour chaque AESI sont disponibles dans le tableau de bord interactif GVDN Observed vs Expected (OE) [12] .
Tableau 3. Ratios OE agrégés par dernière dose, affections neurologiques, période 0–42 jours.
AESI : SGB = syndrome de Guillain-Barré, TRM = myélite transverse, BP = paralysie faciale (de Bell), ADEM = encéphalomyélite aiguë disséminée, FSZ = convulsions fébriles,
GSZ = Crises généralisées.
Vaccins : Pfizer/BioNTech (BNT162b2), Moderna (ARNm-1273) et Oxford/Astra Zeneca/Serum Institute of India (ChAdOx1).
Tableau 4. Ratios OE agrégés par dernière dose, conditions hématologiques, période 0–42 jours.
Vaccines: Pfizer/BioNTech (BNT162b2), Moderna (mRNA-1273), and Oxford/Astra Zeneca/Serum Institute of India (ChAdOx1).
Table 5. Aggregated OE Ratios by last dose, cardiovascular conditions, period 0–42 days.
3.1 . Affections neurologiques
Français Il y a eu une augmentation statistiquement significative des cas de SGB dans les 42 jours suivant une première dose de ChAdOx1 (ratio OE = 2,49 ; IC à 95 % : 2,15, 2,87), indiquant un signal de sécurité prioritaire ( Tableau 3 ). Soixante-seize événements de SGB étaient attendus et 190 événements ont été observés ( Fig. 1 ). Le ratio OE pour l’ADEM dans les 42 jours suivant une première dose d’ARNm-1273 a également atteint le seuil de signification d’un signal de sécurité prioritaire (3,78 ; IC à 95 % : 1,52, 7,78), avec deux événements attendus contre sept événements observés ( Fig. 1 ).
Français Des différences statistiquement significatives ont également été trouvées pour la myélite transverse (ratio OE = 1,91 ; IC à 95 % : 1,22, 2,84) et l’ADEM (ratio OE = 2,23 ; IC à 95 % : 1,15, 3,90) après une première dose de ChAdOx1. La paralysie de Bell avait un ratio OE accru après une première dose de BNT162b2 (1,05 ; IC à 95 % : 1,00, 1,11) et d’ARNm-1273 (1,25 ; IC à 95 % : 1,11, 1,39). Français On a également observé une augmentation des ratios d’OE pour les convulsions fébriles après une première et une deuxième dose d’ARNm-1273 (1,36, IC à 95 % : 1,02, 1,77 et 1,44, IC à 95 % : 1,04, 1,95, respectivement), et pour les convulsions généralisées après une première dose d’ARNm-1273 (1,15, IC à 95 % : 1,10, 1,20) et une quatrième dose de BNT162b2 (1,09, IC à 95 % : 1,04, 1,14). Aucune augmentation des ratios d’OE n’a été identifiée après une troisième dose d’un vaccin. Les résultats concordent avec les ratios d’OE des calendriers homologues ; Cependant, un ratio OE accru pour les crises généralisées après un schéma homologue de quatre doses d’ARNm-1273 (1,33 ; IC à 95 % : 1,07, 1,63) a été identifié ( Fig. 1 ). Ces résultats n’ont pas atteint le seuil d’un signal de sécurité prioritaire après la vaccination.
3.2 . Affections hématologiques
Le rapport OE de CVST était de 3,23 (IC à 95 % : 2,51–4,09) dans les 42 jours suivant une première dose de ChAdOx1, remplissant le seuil d’un signal de sécurité prioritaire ( Tableau 4 ). Au total, 21 événements étaient attendus, tandis que 69 événements ont été observés ( Fig. 2 ).
Français Des ratios OE accrus ont également été identifiés pour la thrombocytopénie après une première dose de ChAdOx1 (1,07 ; IC à 95 % : 1,03, 1,12), BNT162b2 (1,11 ; IC à 95 % : 1,08, 1,14) et mRNA-1273 (1,33 ; IC à 95 % 1,25, 1,42), ainsi qu’après une troisième dose de ChAdOx1 (1,95 ; IC à 95 % : 1,29, 2,84). La thrombocytopénie immunitaire a également démontré des ratios OE accrus après une première dose de ChAdOx1 (1,40 ; IC à 95 % : 1,24, 1,58) et BNT162b2 (1,08 ; IC à 95 % : 1,01, 1,16). Français Les ratios OE d’embolie pulmonaire ont augmenté après les premières doses de ChAdOx1 (1,20 ; IC à 95 % : 1,16, 1,24), BNT162b2 (1,29 ; IC à 95 % : 1,26, 1,32) et mRNA-1273 (1,33, IC à 95 % : 1,26, 1,40), ainsi qu’après une troisième dose de ChAdOx1 (1,88 ; IC à 95 % : 1,32, 2,58). Le ratio OE de CVST était de 1,49 (IC à 95 % : 1,26, 1,75) après une première dose et de 1,25 (IC à 95 % : 1,06, 1,46) après une deuxième dose de BNT162b2. Français Un ratio OE accru pour la tachycardie supraventriculaire a été observé après une première dose de BNT162b2 (1,25 ; IC à 95 % : 1,17, 1,34) et d’ARNm-1273 (1,23 ; IC à 95 % : 1,03, 1,47) ; une deuxième dose d’ARNm-1273 (1,17 ; IC à 95 % : 1,01, 1,36) ; et une quatrième dose de BNT162b2 (1,30, IC à 95 % : 1,06, 1,59) et d’ARNm-1273 (1,53, IC à 95 % : 1,05, 2,16). Ces résultats n’ont pas atteint le seuil d’un signal de sécurité prioritaire après la vaccination.
3.3 . Affections cardiovasculaires
Français Des ratios OE accrus satisfaisant le seuil des signaux de sécurité prioritaires pour la myocardite ont été systématiquement identifiés après une première, une deuxième et une troisième dose de vaccins à ARNm (BNT162b2 et ARNm-1273) ( Tableau 4 ). Le ratio OE le plus élevé a été observé après une première et une deuxième dose d’ARNm-1273 (3,48 ; IC à 95 % : 3,00, 4,01 et 6,10 ; IC à 95 % : 5,52, 6,72, respectivement). Le ratio OE après une troisième dose d’ARNm-1273 était de 2,01 (IC à 95 % : 1,60, 2,49). Le nombre d’événements pour un maximum de quatre doses de calendriers homologues est présenté dans la Fig. 3. Les ratios OE des calendriers homologues correspondent aux ratios OE agrégés. L’OE homologue pour la myocardite après quatre doses du vaccin à ARNm-1273 n’a pas pu être estimé en raison d’un manque d’événements observés.
Français De même, le ratio OE pour la péricardite a atteint le seuil d’un signal de sécurité prioritaire après une première et une quatrième dose d’ARNm-1273, avec des ratios OE de 1,74 (IC à 95 % : 1,54, 1,97) et 2,64 (IC à 95 % : 2,05, 3,35) respectivement. Un ratio accru de 6,91 (IC à 95 % : 3,45, 12,36), atteignant le seuil d’un signal de sécurité prioritaire, a également été observé après une troisième dose de ChAdOx1. Les ratios OE agrégés pour la péricardite ont augmenté après toutes les doses des trois vaccins présentés ( Tableau 4 ). Français Les résultats sont très similaires aux ratios des schémas homologues ( Fig. 3 ), à l’exception du ratio OE de 1,23 (IC à 95 % : 0,45–2,69) après réception de la quatrième dose d’ARNm-1273, qui n’a pas atteint le seuil de signal de sécurité. Le ratio OE homologue après une troisième dose de ChAdOx1 n’a pas été rapporté car seul un petit nombre de troisièmes doses de ChAdOx1 ont été administrées dans les sites d’étude ( Tableau 1 ).
3.4 . Analyse de sensibilité
Français Des analyses secondaires ont été menées pour explorer plus en détail le SGB, l’ADEM, la TVC, la myocardite et la péricardite au niveau du site spécifique. Nous rapportons les ratios d’OE agrégés par dernière dose et site dans la période de 0 à 42 jours après la vaccination dans les tableaux supplémentaires 6 à 10. Il n’a pas été possible de rapporter les résultats pour tous les sites et les résultats de l’étude en raison d’un nombre insuffisant de personnes-années ou de moins de cinq événements observés selon les critères de confidentialité du site. La majorité des signaux de sécurité identifiés suite à des combinaisons spécifiques de marques et de doses de vaccins dans l’analyse principale ont cependant été confirmés par les sites individuels où les données étaient disponibles. L’analyse supplémentaire avec un seuil de personnes-années de 1 000 et incluant d’autres vaccins et doses administrés dans les sites GVDN, a montré un ratio d’OE accru pour certains résultats, par exemple pour les crises généralisées après une première dose du vaccin Gamaleya Research Institute/Sputnik (5,50, IC à 95 % : 2,74, 9,84) ( Tableaux supplémentaires 11 à 13 ).
4. Discussion
multinationale Cette étude de cohorte a été menée dans le cadre unique du GVDN. À ce jour, le nombre d’études de grande envergure, coordonnées systématiquement et portant sur des zones géographiques et des populations diverses, est limité. Cependant, plusieurs études ont déjà évalué les risques liés aux signaux de sécurité identifiés après la vaccination contre la COVID-19, principalement dans des contextes monocentriques. Nous avons étudié l’association entre la vaccination contre la COVID-19 et 13 effets indésirables graves (EIS) comprenant des affections neurologiques, hématologiques et cardiovasculaires sur 10 sites répartis dans huit pays, dont l’Europe, l’Amérique du Nord, l’Amérique du Sud et l’Océanie. Dans cette étude incluant plus de 99 millions de personnes vaccinées contre le SARS-CoV-2, le risque jusqu’à 42 jours après la vaccination était généralement similaire au risque de base pour la majorité des résultats ; cependant, quelques signaux de sécurité potentiels ont été identifiés. Nous avons observé des signaux de sécurité potentiels pour le SGB et la TVC après la première dose de ChAdOx1, sur la base de plus de 12 millions de doses administrées.
Français Dans l’ensemble, les études sur les vaccins à base de vecteurs tels que le ChAdOx1 ont observé une incidence plus élevée de SGB après la vaccination par rapport à l’incidence de base ; alors que la plupart des études sur les vaccins à ARNm , tels que le BNT162b2 et l’ARNm-1273, n’ont pas observé d’augmentation de SGB [15] , [24] , [14] , [25] , [26] , [27] . Atzenhoffer et al. [24] ont signalé un ratio OE élevé > 2,0 pour les vaccins COVID-19 à vecteur adénovirus, dans les pays contribuant à VigiBase, une base de données internationale sur les effets indésirables des médicaments et Patone et al. [27] ont signalé 38 cas excédentaires de SGB pour 10 millions de personnes exposées au cours de la période à risque de 1 à 28 jours suivant la vaccination avec le ChAdOx1 en Angleterre. Les auteurs n’ont pas observé de risque accru chez ceux qui ont reçu le BNT162b2. En revanche, une étude de Li et al. [28] n’ont montré aucun risque accru de SGB pour ChAdOx1, tandis que seule l’infection par le SARS-CoV-2 était associée à un risque plus élevé. Cet écart, comparé aux résultats de Patone et al. [27] , pourrait toutefois s’expliquer par un échantillon plus petit et des mesures de résultats différentes. Globalement, ces données corroborent nos conclusions concernant un signal d’alerte concernant le SGB après la vaccination par ChAdOx1. Bien que rare, cette association a été reconnue par l’OMS, l’Agence européenne des médicaments (EMA) et la Therapeutic Goods Administration (TGA) d’Australie, ce qui a conduit à classer le SGB comme une maladie rare. effet secondaire suite à une exposition à ChAdOx1 [15] , [29] , [30] .
Français Le risque accru de TVC identifié après la vaccination par ChAdOx1 dans cette étude est corroboré par de multiples études. Un ratio OE accru a été observé dans une étude de cohorte nationale au Danemark et en Norvège, avec des taux accrus d’ événements thromboemboliques veineux , y compris la TVC avec un taux excédentaire de 2,5 événements pour 100 000 vaccinations après ChAdOx1 [7] . Sur la base de diverses méthodologies, d’autres études ont également signalé une incidence accrue de TVC après la vaccination [31] , [32] . En fin de compte, ce signal de sécurité rare mais préoccupant a conduit au retrait du vaccin ChAdOx1 des programmes de vaccination contre la COVID-19 ou à la mise en œuvre de restrictions basées sur l’âge dans plusieurs pays [8] .
Il est crucial de reconnaître le seuil de signification des signaux de sécurité prioritaires appliqués dans cette étude (LBCI > 1,5). Ce seuil a été sélectionné sur la base de l’avis d’experts au sein du GVDN et des CDC, afin de se concentrer sur les résultats les plus susceptibles d’être de véritables signaux. Certains événements observés, bien que n’atteignant pas ce seuil, peuvent néanmoins avoir une importance clinique et nécessiter des investigations plus approfondies. Par exemple, un PTI avec un ratio OE > 1,0 et un LBCI de 1,2 après vaccination par ChAdOx1 concorde avec les résultats rapportés dans la littérature comme un signal potentiel. Cette concordance est mise en évidence dans une étude menée à Victoria, en Australie, qui a observé un taux de PTI nettement plus élevé que prévu après vaccination par ChAdOx1 [33] .
Français De plus, nous avons observé des risques significativement plus élevés de myocardite après les première, deuxième et troisième doses de BNT162b2 et d’ARNm-1273 ainsi que de péricardite après la première et la quatrième dose d’ARNm-1273, et la troisième dose de ChAdOx1, dans la période à risque de 0 à 42 jours. Les taux élevés de péricardite après la vaccination par ChAdOx1 identifiés dans cette étude reposent sur un nombre limité de décomptes observés dans la méta-analyse. Le large intervalle de confiance souligne l’incertitude substantielle de la caractérisation de la péricardite comme un signal de sécurité après la vaccination par ChAdOx1. Cependant, notre étude confirme les résultats de cas rares de myocardite et de péricardite précédemment identifiés après la première et la deuxième dose de vaccins à ARNm[21] , [22] , [23] , [34] . Une vaste étude de cohorte portant sur 23,1 millions de résidents de quatre pays nordiques a révélé un risque accru de myocardite chez les jeunes hommes âgés de 16 à 24 ans, sur la base de 4 à 7 événements excédentaires en 28 jours pour 100 000 vaccinés après une deuxième dose de BNT162b2, et entre 9 et 28 pour 100 000 vaccinés après une deuxième dose de mRNA-1273 [22] . De même, des études menées en Colombie-Britannique, au Canada, ont signalé que les cas de myocardite étaient plus élevés chez les personnes ayant reçu une deuxième dose par rapport à une troisième dose, et chez celles ayant reçu une deuxième dose du vaccin à ARNm-1273 par rapport au vaccin BNT162b2 [35] , [36] . Patone et al. [37] ont estimé que les événements de myocardite supplémentaires se situaient entre un et 10 par million de personnes au cours du mois suivant la vaccination, ce qui était nettement inférieur aux 40 événements supplémentaires par million de personnes observés après la période d’infection par le SRAS-CoV-2. Une revue systématique d’Alami et al. [38] a conclu que les personnes vaccinées à l’ARNm étaient deux fois plus susceptibles de développer une myocardite/péricardite que les personnes non vaccinées, avec un rapport de taux de 2,05 (IC à 95 % 1,49–2,82). Compte tenu des preuves, l’OMS a publié des directives actualisées concernant ces signaux de sécurité et la vaccination à ARNm contre la COVID-19, et l’EMA a fourni des mises à jour des informations sur le produit pour les vaccins BNT162b2 et ARNm-1273 [21] , [23] . La TGA ainsi que les CDC continuent de surveiller et d’examiner les données sur la myocardite et la péricardite après la vaccination contre la COVID-19. [39] , [40] .
Français Un autre signal de sécurité potentiel a été identifié pour l’ADEM après la première dose du vaccin à ARNm-1273, avec cinq événements observés de plus que prévu sur la base de 1 035 871 personnes-années et 10,5 millions de doses administrées ; cependant, le nombre de cas de cet événement rare était faible et l’intervalle de confiance large, de sorte que les résultats doivent être interprétés avec prudence et confirmés dans de futures études. Bien que certains rapports de cas aient suggéré une association possible entre la vaccination contre la COVID-19 et l’ADEM, il n’y avait pas de tendance cohérente en termes de vaccin ou de moment après la vaccination, et des études épidémiologiques de plus grande envergure n’ont confirmé aucune association potentielle [41] , [42] , [43] , [44] . De plus, les rapports de cas peuvent faire état d’événements fortuits et n’établissent pas d’association ni n’indiquent de causalité, par conséquent des études observationnelles de plus grande envergure sont nécessaires pour approfondir nos résultats. Pour y remédier, une étude de suivi est actuellement menée au sein du GVDN, se concentrant sur un groupe démographique non inclus dans notre analyse. Sur la base des rapports de cas rares d’ADEM à la base de données européenne des effets indésirables suspectés des médicaments, l’EMA a évalué l’association potentielle d’ADEM après la vaccination avec ChAdOx1 [45] . Frontera et al. [46] ont conclu que les risques d’avoir un événement neurologique après une infection aiguë par le SRAS-CoV-2 étaient jusqu’à 617 fois plus élevés qu’après une vaccination contre la COVID, ce qui suggère que les avantages de la vaccination l’emportent largement sur les risques. Un signal de sécurité pour les crises généralisées a été identifié après la vaccination Gamaleya Research Institute/Sputnik, mais le nombre de vaccinations était relativement faible par rapport aux autres vaccins dans cette étude. D’autres études sont nécessaires pour explorer ce signal de sécurité potentiel.
La réalisation d’une analyse de cohorte dans le contexte multi-pays unique du GVDN s’appuie sur un bassin de données vaste et diversifié. L’agrégation de données provenant de plusieurs pays, portant sur plus de 99 millions de personnes vaccinées, a considérablement augmenté la taille de l’échantillon et la puissance statistique par rapport à de nombreuses études de sécurité antérieures. Cela améliore la capacité à détecter les signaux d’innocuité, notamment pour les effets indésirables extrêmement rares, car la taille plus importante de l’échantillon permet une estimation plus précise des taux observés.
Les résultats basés sur des données provenant d’Europe, d’Amérique du Nord et du Sud, ainsi que d’Océanie, offrent une validité externe plus solide, permettant une généralisation plus large des résultats à un éventail plus large de populations et de structures de santé participant au programme mondial de vaccination contre la COVID-19. De plus, les analyses multi-pays facilitent les comparaisons entre des pays ayant des stratégies de vaccination, des caractéristiques démographiques et des systèmes de santé différents, ce qui permet de mieux comprendre l’influence de ces facteurs sur les profils de sécurité vaccinale. Les données utilisées dans notre analyse proviennent de plusieurs bases de données, notamment des bases de données de santé, des registres nationaux de vaccination et des tableaux de bord de vaccination, ce qui permet d’identifier des signaux potentiels de sécurité provenant de diverses sources.
Les résultats de notre étude doivent toutefois être interprétés en tenant compte de multiples limites. Nos analyses impliquent intrinsèquement une hétérogénéité dans la collecte, la qualité et les normes de déclaration des données entre les pays. Ces différences dans les infrastructures de santé et les systèmes de surveillance peuvent introduire des biais et affecter la comparabilité des résultats. Les sites participants des huit pays ont mis en œuvre des stratégies de vaccination variées, notamment en ce qui concerne les types de vaccins, les schémas posologiques et la priorisation des vaccinés. De plus, les analyses multipays sont sensibles aux facteurs de confusion démographiques, tels que les différences dans les conditions de santé préexistantes, les facteurs génétiques , les profils ethniques et les comportements, qu’il n’a pas été possible de prendre en compte dans notre analyse. Nous considérons que notre approche est adaptée à une application à de grands ensembles de données représentant des populations moyennes. Cependant, les taux historiques spécifiques à l’âge et au sexe qui ne sont pas ajustés pour des facteurs tels que les maladies antérieures peuvent ne pas fournir une comparaison pertinente, par exemple, dans les premières phases d’une campagne de vaccination où les personnes présentant des comorbidités ont été vaccinées avant les autres groupes de population.
La sous-déclaration potentielle entre les pays pourrait avoir conduit à une sous-estimation de l’importance des signaux potentiels de sécurité. Il est important de reconnaître le risque de faux négatifs , en particulier lors de la détection d’associations avec des intervalles de confiance inférieurs à 1,5 qui maintiennent une signification statistique. Les signaux de sécurité identifiés dans cette étude doivent être évalués en fonction de leur rareté, de leur gravité et de leur pertinence clinique. De plus, les évaluations globales du rapport bénéfice/risque de la vaccination doivent tenir compte du risque associé à l’infection, car plusieurs études ont démontré un risque plus élevé de développer les événements étudiés, tels que le syndrome de Guillain-Barré (SGB), la myocardite ou l’EMA, après une infection par le SARS-CoV-2 que la vaccination. Enfin, l’utilisation des codes CIM-10 est soumise à des considérations de spécificité et de sensibilité, et son application peut varier selon les pays.
5. Conclusion
Les analyses comparatives observées/attendues dans le contexte multi-pays du GVDN et du projet GCoVS offrent un ensemble de données plus vaste et plus diversifié, une meilleure généralisabilité et une puissance statistique accrue par rapport aux études monocentriques ou régionales. Elles présentent également des défis liés à l’hétérogénéité des données, aux facteurs de confusion démographiques et aux variations des stratégies de vaccination et des systèmes de déclaration. L’implication de chercheurs et de sources de données provenant de diverses régions du monde favorise l’inclusion, réduit les biais potentiels et encourage la collaboration dans la poursuite d’un objectif commun de santé publique. Bien que notre étude ait confirmé des signaux d’innocuité rares identifiés suite à la vaccination contre la COVID-19 et apporté des preuves sur plusieurs autres résultats importants, des investigations complémentaires sont nécessaires pour confirmer les associations et évaluer la signification clinique . Ceci pourrait être abordé en menant des études d’association spécifiques à des résultats individuels en appliquant des méthodologies telles que les séries de cas auto-contrôlées (SCCS) pour valider les associations. [6] .
Clause de non-responsabilité
Toutes les analyses, conclusions, opinions et déclarations sont celles des auteurs et ne représentent pas nécessairement le point de vue officiel des CDC/HHS ou du gouvernement américain, ni leur approbation. Pour plus d’informations, veuillez consulter cdc.gov.
.
Certaines parties de ce document sont basées sur des données et/ou des informations compilées et fournies par l’Institut canadien d’information sur la santé et le ministère de la Santé de l’Ontario. Les analyses, conclusions, opinions et déclarations exprimées ici sont celles des auteurs et ne reflètent pas celles des sources de financement ou de données ; aucune approbation n’est prévue ni ne doit être déduite. Certaines parties de ce document sont basées sur des données et/ou des informations fournies par le ministère de la Santé de la Colombie-Britannique. Toutes les inférences, opinions et conclusions tirées de ce manuscrit sont celles des auteurs et ne reflètent pas les opinions ou les politiques du ou des responsables des données.
Déclaration de financement
Le projet GCoVS est soutenu par les Centers for Disease Control and Prevention (CDC) du Département de la Santé et des Services Sociaux des États-Unis (HHS) dans le cadre d’une aide financière d’un montant total de 10 108 491 USD, financée à 100 % par le CDC/HHS .
Le site ontarien contribuant à cette étude a été soutenu par Santé publique Ontario et par l’ ICES , qui est financé par une subvention annuelle du ministère de la Santé de l’Ontario de l’ . JCK est soutenu par une bourse de clinicien-chercheur du département de médecine familiale et communautaire Université de Toronto .
Déclaration de contribution d’auteur Crédit
K. Faksova : Visualisation, Rédaction – brouillon original, Rédaction – révision et édition. D. Walsh : Conservation des données, Analyse formelle, Investigation, Méthodologie, Logiciel, Validation, Conceptualisation, Rédaction – révision et édition, Visualisation. Y. Jiang : Conceptualisation, Conservation des données, Analyse formelle, Investigation, Méthodologie, Logiciel, Supervision, Validation, Visualisation, Rédaction – révision et édition. J. Griffin : Conceptualisation, Rédaction – révision et édition, Méthodologie. A. Phillips : Conceptualisation, Méthodologie, Rédaction – révision et édition, Investigation, Validation. A. Gentile : Conservation des données, Investigation, Supervision, Validation. JC Kwong : Conservation des données, Supervision, Validation, Rédaction – révision et édition, Investigation, Méthodologie. K. Macartney : Conservation des données, Supervision, Validation, Rédaction – révision et édition, Investigation, Méthodologie. M. Naus : Conservation des données, Supervision, Validation, Investigation, Méthodologie. Z. Grange : Conservation des données, Supervision, Validation, Conceptualisation, Investigation, Méthodologie. S. Escolano : Conservation des données, Supervision, Validation, Investigation, Méthodologie, Rédaction – révision et édition. G. Sepulveda : Conservation des données, Analyse formelle, Logiciel, Validation. A. Shetty : Conservation des données, Validation, Investigation, Méthodologie. A. Pillsbury : Conservation des données, Validation, Investigation, Méthodologie, Rédaction – révision et édition. C. Sullivan : Conservation des données, Validation, Investigation, Méthodologie, Rédaction – révision et édition. Z. Naveed : Conservation des données, Validation, Investigation, Méthodologie, Rédaction – révision et édition. NZ Janjua : Conservation des données, Rédaction – révision et édition. N. Giglio : Conservation des données, Investigation, Méthodologie, Validation. J. Perälä : Conservation des données, Investigation, Méthodologie, Validation. S. Nasreen : Conceptualisation, Conservation des données, Validation, Rédaction – révision et édition. H. Gidding : Conceptualisation, Validation, Rédaction – révision et édition, Investigation, Méthodologie. P. Hovi : Conceptualisation, Validation, Rédaction (révision et correction), Investigation, Méthodologie. T. Vo : Conceptualisation, Validation, Analyse formelle, Investigation, Méthodologie, Rédaction – révision et édition. F. Cui : Conceptualisation, Investigation, Méthodologie, Validation. L. Deng : Conceptualisation, Investigation, Méthodologie, Validation, Rédaction – révision et édition. L. Cullen : Conceptualisation, Investigation, Méthodologie, Validation, Rédaction – révision et édition. M. Artama : Conceptualisation, Investigation, Méthodologie, Validation, Rédaction – révision et édition. H. Lu : Conservation des données, Analyse formelle, Logiciel, Validation. HJ Clothier : Conceptualisation, Méthodologie, Validation, Rédaction – révision et édition, Conservation des données, Analyse formelle, Administration de projet. K. Batty : Conceptualisation, Méthodologie, Administration de projet, Validation, Rédaction – révision et édition. J. Paynter : Conceptualisation, Méthodologie, Supervision, Conservation des données, Analyse formelle, Rédaction – révision et édition. H. Petousis-Harris : Conceptualisation, Acquisition de financement, Enquête, Méthodologie, Ressources, Supervision, Rédaction – révision et édition, Administration de projet. J. Buttery : Conceptualisation, Acquisition de financement, Investigation, Méthodologie, Ressources, Supervision, Validation, Rédaction – révision et édition, Administration de projet. S. Black : Conceptualisation, Acquisition de financement, Investigation, Méthodologie, Administration de projet, Ressources, Supervision, Validation, Rédaction – révision et édition. A. Hviid : Conceptualisation, Investigation, Méthodologie, Supervision, Validation, Rédaction – révision et édition.
Déclaration d’intérêts concurrents
Français Les auteurs déclarent les intérêts financiers/relations personnelles suivants qui peuvent être considérés comme des intérêts concurrents potentiels : Jeffrey C. Kwong rapporte que le soutien financier a été fourni par les Centers for Disease Control and Prevention. Naveed Z. Janjua rapporte que le soutien financier a été fourni par les Centers for Disease Control and Prevention. Anders Hviid rapporte que le soutien financier a été fourni par le Global Vaccine Data Network. Helen Petousis-Harris rapporte que le soutien financier a été fourni par le ministère de la Santé de la Nouvelle-Zélande. Steven Black rapporte une relation avec GSK qui comprend : des services de consultation ou de conseil. Jeffrey C. Kwong rapporte une relation avec les Instituts de recherche en santé du Canada qui comprend : des subventions de financement. Jeffrey C. Kwong rapporte une relation avec l’Agence de la santé publique du Canada qui comprend : des subventions de financement. Naveed Z. Janjua rapporte une relation avec AbbVie Inc qui comprend : des services de consultation ou de conseil et des honoraires de conférencier et de conférence. Naveed Z. Janjua rapporte une relation avec Gilead Sciences Inc qui comprend : des honoraires de conférencier et de conférence. Anders Hviid rapporte une relation avec l’Independent Research Fund Denmark qui comprend : des subventions de financement. Anders Hviid déclare entretenir des relations avec la Fondation Lundbeck, notamment des subventions de financement. Anders Hviid déclare entretenir des relations avec la Fondation Novo Nordisk, notamment des subventions de financement. Anders Hviid déclare entretenir des relations avec VAC4EU, notamment des activités de conseil ou de consultation. L’Institut finlandais pour la santé et le bien-être (THL) mène des partenariats public-privé avec des fabricants de vaccins et a reçu des financements de recherche de Sanofi Inc. Petteri Hovi a participé à ces études en tant que chercheur, mais n’a perçu aucune rémunération personnelle. Helen Petousis-Harris a siégé à des comités consultatifs d’experts et a donné des conférences pour Pfizer et GSK. Elle a également reçu des financements de recherche de GSK. Elle n’a perçu aucun honoraire personnel. Si d’autres auteurs sont présents, ils déclarent n’avoir aucun intérêt financier concurrent ni aucune relation personnelle connus qui auraient pu influencer les travaux présentés dans cet article.
Remerciements
Le protocole d’étude des analyses observées et attendues des événements indésirables du vaccin contre la COVID-19 d’intérêt particulier a été développé par le groupe de travail observé et attendu dirigé par Anders Hviid. a, b Les membres du groupe de travail étaient Nelson Aguirre Duarte c , Miia Artama d , Karin Batty et , Steven Black c, f , Hannah Chisholm c , Hazel Clothier g, h, i , Fuqiang Cui j , Lucy Deng k Lucy Cullen l , Heather Gidding k, m, n , Petteri Hovi le , Yannan Jiang c Janine Paynter c , Helen Petousis-Harris c Anastasia Phillips k , John Sluyter c , Thuan Vo faire , et Daniel Walsh c , Éric Weintraub p .
Affiliations aux groupes de travail
a. Statens Serum Institut, Danemark ; b. Université de Copenhague, Danemark ; c. Université d’Auckland, Nouvelle-Zélande ; d. Université de Tampere, Finlande ; e. Auckland UniServices Limited à l’Université d’Auckland, Nouvelle-Zélande ; f. Université de Cincinnati et Hôpital pour enfants, États-Unis ; g. Département de la santé de Victoria, Australie ; h. Institut de recherche pour enfants Murdoch, Australie ; i. Centre d’analyse de la santé, Campus pour enfants de Melbourne, Australie ; j. Université de Pékin, Pékin, Chine ; k. Centre national de recherche sur l’immunisation et de surveillance des maladies évitables par la vaccination, Australie ; l. Public Health Scotland, Glasgow, Écosse, Royaume-Uni ; m. Institut Kolling, District sanitaire local de Northern Sydney, Australie ; n. École clinique du Nord de l’Université de Sydney, Australie ; o. Institut finlandais pour la santé et le bien-être, Finlande ; p. Vaccine Safety Datalink, Centres pour le contrôle et la prévention des maladies.
Les personnes suivantes ont contribué en tant que responsables de site du GVDN : Anders Hviid (Danemark) ; Angela Gentile (Argentine) ; Sylvie Escolano (France) ; Eero Poukka (Finlande) ; Jeffrey C. Kwong (Ontario, Canada) ; Kristine Macartney (Nouvelle-Galles du Sud, Australie) ; Jim Buttery (Victoria, Australie) ; Monika Naus (Colombie-Britannique, Canada) ; Zoe Grange (Écosse) ; et Helen Petousis-Harris (Nouvelle-Zélande).
Les personnes suivantes ont contribué en tant qu’enquêteurs du site GVDN : Gonzalo Sepulveda et Aishwarya Shetty (Victoria, Australie) ; Alexis Pillsbury (Nouvelle-Galles du Sud, Australie) ; Christopher Sullivan (Écosse) ; Naveed Zaeema (Colombie-Britannique, Canada) ; Norberto Giglio (Argentine) ; Jori Perälä (Finlande) ; Sharifa Nasreen (Ontario, Canada) ; Han Lu (Nouvelle-Zélande).
Plateforme de sécurité pour les vaccins émergents S. D2.3 Liste prioritaire des effets indésirables présentant un intérêt particulier : COVID-19. Brighton Collaboration ; 2020. Disponible sur : https://zenodo.org/records/6656179#.Y-0yxuyZOnN
MDX van der Boom , R. van Eekeren , FPAM van Hunsel
Analyse des observations supérieures aux attentes comme méthode supplémentaire de détection des signaux de pharmacovigilance dans les rapports spontanés d’événements indésirables à grande échelle
Évaluation de l’efficacité et de la puissance statistique des séries de cas autocontrôlées et des modèles d’intervalles de risque autocontrôlés dans la sécurité des vaccins
A. Pottegård , LC Lund , Ø. Karlstad , J. Dahl , M. Andersen , J. Hallas , et al.
Événements artériels, thromboembolie veineuse, thrombocytopénie et saignements après vaccination avec Oxford-AstraZeneca ChAdOx1-S au Danemark et en Norvège : étude de cohorte basée sur la population
Agence européenne des médicaments. Rapport d’évaluation des signaux sur les événements emboliques et thrombotiques (SMQ) avec le vaccin contre la COVID-19 (ChAdOx1-S [recombinant])-Vaxzevria (anciennement vaccin AstraZeneca contre la COVID-19) (autres vaccins viraux). 2021.
Réseau mondial de données sur les vaccins. Auckland (NZ) : Réseau mondial de données sur les vaccins ; 2023 [consulté le 13 octobre 2023]. Disponible sur : https://www.globalvaccinedatanetwork.org/
Réseau mondial de données sur les vaccins. GVDN : Tableau de bord des données observées et attendues (OE). Auckland (NZ) : Réseau mondial de données sur les vaccins ; 2023 [mis à jour le 8 décembre 2023 ; cité le 13 décembre 2023]. Disponible sur : www.globalvaccinedatanetwork.org/Data-Dashboards
A. Phillips , Y. Jiang , D. Walsh , N. Andrews , M. Artama , H. Clothier , et al.
Taux de référence des événements indésirables présentant un intérêt particulier pour les vaccins contre la COVID-19 : une analyse multinationale du Global Vaccine Data Network (GVDN)
Organisation mondiale de la Santé. Déclaration du sous-comité COVID-19 du Comité consultatif mondial de l’OMS sur la sécurité des vaccins (GACVS) concernant les cas de syndrome de Guillain-Barré (SGB) signalés après l’administration de vaccins à vecteur adénoviral contre la COVID-19. nd https://www.who.int/news/item/26-07-2021-statement-of-the-who-gacvs-covid-19-subcommittee-on-gbs
E. Burn , X. Li , K. Kostka , H. M. Stewart , C. Reich , S. Seager , et al.
Taux de base de cinq syndromes de thrombose avec thrombocytopénie présentant un intérêt particulier pour la surveillance de la sécurité des vaccins contre la COVID-19 : incidence entre 2017 et 2019 et profils de patients de 38,6 millions de personnes dans six pays européens
A. Hviid , J.V. Hansen , E.M. Thiesson , J. Wohlfahrt
Association de la vaccination contre la COVID-19 par AZD1222 et BNT162b2 avec des événements thromboemboliques et thrombocytopéniques chez le personnel de première ligne : une étude de cohorte rétrospective
JW Gargano , M. Wallace , SC Hadler , G. Langley , JR Su , ME Oster , et al.
Utilisation du vaccin à ARNm contre la COVID-19 après des cas de myocardite chez des personnes vaccinées : mise à jour du comité consultatif sur les pratiques de vaccination — États-Unis, juin 2021
M. Otero-Losada , N. Petrovsky , A. Alami , J.A. Crispo , D. Mattison , F. Capani , et al.
Analyse de disproportionnalité des effets indésirables neurologiques et psychiatriques des vaccins ChAdOx1 (Oxford-AstraZeneca) et BNT162b2 (Pfizer-BioNTech) contre la COVID-19 au Royaume-Uni
JL Walker , A. Schultze , J. Tazare , A. Tamborska , B. Singh , K. Donegan et al.
Sécurité de la vaccination contre la COVID-19 et événements neurologiques aigus : une série de cas autocontrôlés en Angleterre utilisant la plateforme OpenSAFELY
X. Li , B. Raventós , E. Roel , A. Pistillo , E. Martinez-Hernandez , A. Delmestri , et al.
Association entre la vaccination contre la covid-19, l’infection par le SRAS-CoV-2 et le risque d’événements neurologiques à médiation immunitaire : analyse de cohorte basée sur la population et analyse de séries de cas autocontrôlées
Andrews NJ, Stowe J, Ramsay ME, Miller E. Risque d’événements thrombotiques veineux et de thrombocytopénie sur des périodes séquentielles après les vaccins ChAdOx1 et BNT162b2 contre la COVID-19 : une étude de cohorte nationale en Angleterre. Lancet Reg Heal Eur 2022 ; 13. doi : 10.1016/J.LANEPE.2021.100260.
SA Buchan , CY Seo , C. Johnson , S. Alley , JC Kwong , S. Nasreen , et al.
Épidémiologie de la myocardite et de la péricardite après vaccination par ARNm, selon le produit vaccinal, le calendrier et l’intervalle interdose chez les adolescents et les adultes en Ontario, au Canada
A. Alami , D. Krewski , N. Farhat , D. Mattison , K. Wilson , CA Gravel , et al.
Risque de myocardite et de péricardite dans les populations vaccinées et non vaccinées contre la COVID-19 par ARNm : revue systématique et méta-analyse
J.A. Frontera , A.A. Tamborska , M.F. Doheim , D. Garcia-Azorin , H. Gezegen , A. Guekht , et al.
Événements neurologiques signalés après l’administration des vaccins contre la COVID-19 : une analyse du système de déclaration des événements indésirables liés aux vaccins
Demande de traitement humain et de libération immédiate du Dr Reiner Füellmich. La communauté internationale des signataires de cette déclaration de soutien exige la libération immédiate du Dr Reiner Füellmich, avocat et principal critique des mesures COVID-19, emprisonné depuis octobre 2023.
Le Dr Reiner Füelmich est en détention provisoire depuis octobre 2023. Il a été arrêté dans des circonstances douteuses à l’aéroport de Francfort le 13 octobre 2023.
L’affaire du Dr Füellmich a soulevé de sérieuses inquiétudes quant à la légalité de sa détention et à l’intégrité du processus judiciaire. Des 18 chefs d’accusation initiaux contre Füellmich, il n’en reste plus qu’un, relatif à des prêts personnels.
En vertu du droit allemand, la durée maximale de la détention provisoire est de six mois, comme décrit dans l’article 121, paragraphe 1, du Code de procédure pénale allemand (StPO). « Les raisons particulières ou importantes justifiant une prolongation de la détention provisoire au-delà de six mois ne sont pas claires. » Cette réclamation souligne la nécessité urgente de réévaluer la détention continue du Dr Fuel me.
Le Dr Füellmich est actuellement détenu à la prison de Rosdorf près de Göttingen, où il est détenu dans des conditions difficiles et isolées. Il est séparé des autres prisonniers, n’a droit qu’à un temps d’isolement dans la cour et sa communication avec le monde extérieur est limitée à seulement trois heures de visites privées par mois.
Cet environnement punitif soulève d’autres questions quant au traitement des personnes en détention provisoire, notamment compte tenu du manque de preuves substantielles justifiant de telles mesures.
Le 11 juin, Reiner Füellmich a de nouveau été placé à l’isolement, un statut qu’il continue de maintenir. Cet isolement signifie qu’il lui est interdit d’avoir des contacts avec d’autres détenus. Les autorités ont justifié cette mesure extrême en affirmant que Füellmich avait fourni des conseils juridiques à ses codétenus, une situation jugée inacceptable par les responsables de sa détention. Füellmich doit manger de manière isolée et n’a droit qu’à une heure par jour pour des activités de plein air, qu’il doit également passer dans une solitude totale. Il n’a pas accès à la salle de sport et ne peut utiliser le téléphone qu’une fois que les autres détenus sont rentrés dans leurs cellules. Ce régime strict souligne la gravité de son internement et des restrictions qui lui sont imposées.
Les accusations portées contre le Dr Füellmich incluent le détournement de fonds, mais de nombreux observateurs, y compris sa défense, soutiennent que ce procès va au-delà des procédures judiciaires ordinaires et est devenu une tentative politiquement motivée de faire taire un éminent critique des mesures COVID-19. Le procès a révélé des changements inquiétants dans les paramètres juridiques, compliquant davantage l’affaire et portant atteinte aux principes de justice.
À la lumière de ces graves allégations et du mépris apparent de la procédure régulière, les signataires de cette pétition demandent la libération immédiate du Dr Reiner Füellmich.
Cette situation ne touche pas seulement un individu, mais constitue également un rappel brutal du potentiel d’influence politique qui peut pénétrer le système judiciaire, sapant ainsi les fondements de la justice et de l’équité.
Alors que le mouvement international pour la liberté observe attentivement la situation, il est essentiel que justice soit faite et que le Dr Füellmich reçoive la liberté qu’il mérite – une liberté qui est essentielle non seulement pour lui, mais aussi pour l’intégrité du système judiciaire lui-même.
Les signatures de cette pétition seront remises à Reiner Fuellmich (ou à son avocat) et au tribunal pour montrer que Reiner Fuellmich n’est pas seul.
Cette remise se fera sur papier (et non numériquement) et ne comportera que les initiales et le nom de famille et sans adresse e-mail.
Un article de Laurent Mucchielli qui s’efforce de synthétiser l’état de la connaissance scientifique sur la criminalité des industries du médicament, à partir des sources américaines et françaises.
Jean-Marc SABATIER est directeur de recherche au CNRS, docteur en biologie cellulaire et microbiologie, habilité à diriger des recherches en biochimie, est un spécialiste des vaccins et thérapies à base de protéines modifiées.
Contexte : Le développement rapide des vaccins contre la COVID-19, combiné à un nombre élevé de rapports d’effets indésirables, a suscité des inquiétudes quant aux mécanismes possibles de lésions, notamment la distribution systémique des nanoparticules lipidiques (LNP) et de l’ARNm, les lésions tissulaires associées à la protéine Spike, la thrombogénicité, le dysfonctionnement du système immunitaire et la cancérogénicité. L’objectif de cette revue systématique est d’étudier les liens de causalité possibles entre l’administration du vaccin contre la COVID-19 et le décès, en utilisant des autopsies et des analyses post-mortem. Méthodes : Nous avons recherché dans PubMed et ScienceDirect tous les rapports d’autopsie et d’autopsie restreinte d’organes publiés concernant la vaccination contre la COVID-19 jusqu’au 18 mai. ème , 2023. Toutes les études d’autopsie et d’autopsie restreinte à des organes incluant la vaccination contre la COVID-19 comme exposition antérieure ont été incluses. L’état des connaissances ayant progressé depuis les publications originales, trois médecins ont examiné chaque cas de manière indépendante et déterminé si la vaccination contre la COVID-19 était la cause directe ou avait contribué de manière significative au décès. Résultats : Nous avons initialement identifié 678 études et, après sélection de nos critères d’inclusion, nous avons inclus 44 articles contenant 325 cas d’autopsie et un cas d’autopsie restreinte à un organe (cœur). L’âge moyen du décès était de 70,4 ans. Le système organique le plus impliqué parmi les cas était le système cardiovasculaire (49 %), suivi par le système hématologique (17 %), le système respiratoire (11 %) et plusieurs systèmes organiques (7 %). Trois systèmes organiques ou plus étaient touchés dans 21 cas. Le délai moyen entre la vaccination et le décès était de 14,3 jours. La plupart des décès sont survenus dans la semaine suivant la dernière administration du vaccin. Au total, 240 décès (73,9 %) ont été jugés de manière indépendante comme étant directement dus ou significativement contribués par la vaccination contre la COVID-19, dont les principales causes de décès comprennent la mort cardiaque subite (35 %), l’embolie pulmonaire (12,5 %), l’infarctus du myocarde (12 %), le VITT (7,9 %), la myocardite (7,1 %), le syndrome inflammatoire multisystémique (4,6 %) et l’hémorragie cérébrale (3,8 %). Conclusions : La concordance observée entre les cas de cette revue et les mécanismes connus de lésions et de décès liés aux vaccins contre la COVID-19, ainsi que la confirmation de l’autopsie par un médecin, suggèrent une forte probabilité d’un lien de causalité entre les vaccins contre la COVID-19 et le décès. Une enquête plus approfondie est nécessaire de toute urgence afin de clarifier nos conclusions.
Introduction
Au 31 mai St En 2023, le SRAS-CoV-2 a infecté environ 767 364 883 personnes dans le monde, entraînant 6 938 353 décès selon l’Organisation mondiale de la santé (OMS) [1]. Ces chiffres officiels sont probablement exagérés en raison de l’utilisation généralisée de la RT-PCR quantitative (RT-qPCR) avec des seuils de cycles arbitrairement élevés comme base du diagnostic de la COVID-19 [2]. En réponse directe à cette catastrophe mondiale, les gouvernements ont adopté une approche coordonnée pour limiter le nombre de cas et la mortalité en combinant des interventions non pharmaceutiques (INP) et de nouvelles plateformes vaccinales basées sur les gènes. Les premières doses de vaccin ont été administrées moins de 11 mois après l’identification de la séquence génétique du SRAS-CoV-2 (aux États-Unis, dans le cadre de l’initiative Operation Warp Speed), ce qui a représenté le développement vaccinal le plus rapide de l’histoire, avec des garanties limitées de sécurité à long terme [3]. Au 31 mai St En 2023, environ 69 % de la population mondiale a reçu au moins une dose d’un vaccin contre la COVID-19 [1].
Français Certaines des plateformes vaccinales COVID-19 utilisées comprennent le virus inactivé (Sinovac – CoronaVac), la sous-unité protéique (Novavax – NVX-CoV2373), le vecteur viral (AstraZeneca – ChAdOx1 nCoV-19, Johnson & Johnson – Ad26.COV2.S) et l’ARN messager (Pfizer-BioNTech – BNT162b2, Moderna – ARNm-1273)[4]. Tous utilisent des mécanismes qui peuvent provoquer des effets indésirables graves ; la plupart impliquent la synthèse incontrôlée de la glycoprotéine Spike comme base de la réponse immunologique. La protéine Spike circulante est le mécanisme délétère probable par lequel les vaccins COVID-19 produisent des effets indésirables [5-9,12,13]. La protéine Spike et/ou les sous-unités/fragments peptidiques peuvent déclencher la dégradation du récepteur ACE2 et la déstabilisation du système rénine-angiotensine (SRA), entraînant une thrombose sévère [5]. Français La protéine Spike active les plaquettes, provoque des lésions endothéliales et favorise directement la thrombose [6]. De plus, les cellules du système immunitaire qui absorbent les nanoparticules lipidiques (LNP) des vaccins contre la COVID-19 peuvent ensuite distribuer systémiquement la protéine Spike et les microARN via les exosomes, ce qui peut entraîner de graves conséquences inflammatoires [6]. De plus, le contrôle du cancer à long terme peut être compromis chez les personnes injectées avec des vaccins à ARNm contre la COVID-19 en raison du dérèglement du facteur de régulation de l’interféron (IRF) et du gène suppresseur de tumeur [6]. De plus, un lien de causalité possible entre les vaccins contre la COVID-19 et diverses maladies a été établi, notamment les troubles neurologiques, la myocardite, les déficits plaquettaires sanguins, les maladies du foie, l’affaiblissement de l’adaptabilité immunitaire et le développement du cancer [6]. Ces résultats sont corroborés par la découverte que la vaccination récurrente contre la COVID-19 avec des vaccins génétiques peut déclencher des niveaux anormalement élevés d’anticorps IgG4, ce qui peut entraîner un dérèglement du système immunitaire et contribuer à l’émergence de maladies auto-immunes, de myocardite et de croissance cancéreuse [7].
Français Les effets neurotoxiques de la protéine Spike peuvent causer ou contribuer au syndrome post-COVID, notamment des céphalées, des acouphènes, un dysfonctionnement autonome et une neuropathie des petites fibres [8]. Spécifique à l’administration de vaccins à vecteur viral contre la COVID-19 (AstraZeneca ; Johnson et Johnson), un nouveau syndrome clinique appelé thrombocytopénie thrombotique immunitaire induite par le vaccin (TTIV) a été identifié en 2021 et caractérisé par le développement de thromboses à des sites corporels atypiques associé à une thrombocytopénie sévère après la vaccination [10]. La pathogenèse de cet effet secondaire potentiellement mortel est actuellement inconnue, bien qu’il ait été suggéré que la TTIV soit causée par des anticorps post-vaccination contre le facteur plaquettaire 4 (PF4) déclenchant une activation plaquettaire importante [10]. Les vaccins à base d’ARNm provoquent rarement une TTIV, mais ils sont associés à une myocardite, ou inflammation du myocarde [11]. Français Les mécanismes de développement de la myocardite après la vaccination contre la COVID-19 ne sont pas clairs, mais on a émis l’hypothèse qu’elle pourrait être causée par le mimétisme moléculaire de la protéine Spike et des auto-antigènes, la réponse immunitaire à l’ARNm et une expression dérégulée des cytokines [11]. Chez les adolescents et les jeunes adultes diagnostiqués avec une myocardite post-vaccinale à ARNm, la protéine Spike libre a été détectée dans le sang tandis que les témoins vaccinés n’avaient pas de protéine Spike circulante [12]. Il a été démontré que les séquences d’ARNm du vaccin contre le SARS-CoV-2 Spike peuvent circuler dans le sang jusqu’à 28 jours après la vaccination [13]. Cependant, les auteurs n’ont pas mesuré l’ARNm du vaccin au-delà de 28 jours. Ces données indiquent que des événements indésirables peuvent survenir pendant une période inconnue après la vaccination, la protéine Spike jouant un rôle étiologique potentiel important.
Parry et al. ont démontré la distribution systémique des LNP contenant de l’ARNm après l’administration du vaccin chez le rat, concluant que les LNP atteignaient leur concentration la plus élevée au site d’injection, suivi du foie, de la rate, des glandes surrénales, des ovaires (femelles) et de la moelle osseuse (fémur) sur 48 heures [14]. De plus, des LNP ont été détectés dans le cerveau, le cœur, les yeux, les poumons, les reins, la vessie, l’intestin grêle, l’estomac, les testicules (mâles), la prostate (mâles), l’utérus (femelles), la thyroïde, la moelle épinière et le sang [14]. Ces données de biodistribution suggèrent que la protéine Spike pourrait être exprimée dans les cellules de nombreux systèmes d’organes vitaux, soulevant des inquiétudes importantes quant au profil de sécurité des vaccins contre la COVID-19. Compte tenu des syndromes de vaccination identifiés et de leurs mécanismes possibles, la fréquence des rapports d’événements indésirables devrait être élevée, en particulier compte tenu du grand nombre de doses de vaccin administrées dans le monde.
Jusqu’au 31 mai St Français En 2024, le Vaccine Adverse Event Reporting System (VAERS) contenait 1 640 416 rapports d’événements indésirables associés aux vaccins contre la COVID-19, dont 37 647 décès, 28 445 myocardites et péricardites, 21 741 crises cardiaques et 9 218 rapports de thrombocytopénie [15]. Si la vaccination contre la COVID-19 est effectivement un déterminant du nombre alarmant de décès signalés, les implications pourraient être immenses, notamment : le retrait complet de tous les vaccins contre la COVID-19 du marché mondial, la suspension de tous les mandats et passeports vaccinaux contre la COVID-19 restants, la perte de confiance du public dans le gouvernement et les institutions médicales, des enquêtes et des investigations sur la censure, le silence et la persécution des médecins et des scientifiques qui ont soulevé ces préoccupations, et une indemnisation pour ceux qui ont été lésés à la suite de l’administration des vaccins contre la COVID-19. Cependant, il n’est pas possible d’utiliser uniquement les données du VAERS pour établir un lien direct entre la vaccination contre la COVID-19 et le décès en raison de nombreuses limitations et facteurs de confusion.
En 2021, Walach et al. ont indiqué que chaque décès après la vaccination contre la COVID-19 devrait faire l’objet d’une autopsie afin d’étudier les mécanismes de préjudice [16]. Les autopsies sont l’un des outils de diagnostic les plus puissants en médecine pour établir la cause du décès et clarifier la physiopathologie de la maladie [17]. Les vaccins contre la COVID-19, avec des mécanismes plausibles de lésion du corps humain et un nombre substantiel de rapports d’événements indésirables, représentent une exposition qui peut être liée de manière causale au décès dans certains cas. L’objectif de cette revue systématique est d’étudier les liens de causalité possibles entre l’administration du vaccin contre la COVID-19 et le décès à l’aide d’analyses post-mortem, y compris des autopsies.
Méthodes
Sources de données et stratégie de recherche
Nous avons effectué une revue systématique de tous les rapports d’autopsie et d’autopsie restreinte d’organe publiés concernant la vaccination contre la COVID-19 jusqu’au 18 mai. ème , 2023. Nous avons utilisé la déclaration PRISMA (Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols) [18] afin de limiter les biais de sélection des études et de garantir la transparence et l’exhaustivité des rapports. Les bases de données suivantes ont été utilisées : PubMed et ScienceDirect. Les mots-clés suivants ont été utilisés : « Vaccin contre la COVID-19 », « Vaccin contre le SARS-CoV-2 » ou « Vaccination contre la COVID » et « Post-mortem » ou « Autopsie ». Toutes les combinaisons de mots-clés possibles ont été recherchées manuellement. La recherche n’était limitée à aucune langue. Toutes les études sélectionnées ont été examinées pour la littérature pertinente contenue dans leurs références.
Critères d’éligibilité et processus de sélection
Tous les articles originaux, essais cliniques randomisés, rapports de cas et séries de cas contenant des résultats d’autopsie ou d’autopsie restreinte à un organe (analyse macroscopique et histologique d’un seul organe) avec des vaccins contre la COVID-19 comme exposition antérieure ont été inclus. Les articles de synthèse, les revues systématiques, les méta-analyses, les articles sans résultats d’autopsie ou d’autopsie restreinte à un organe, les études non humaines et les articles sans statut vaccinal COVID-19 déclaré ont été exclus. Les conclusions des auteurs n’ont pas été prises en compte pour l’inclusion de l’étude afin d’éviter tout biais. Deux auteurs (NH et PAM) ont examiné indépendamment le texte intégral de toutes les études récupérées afin d’évaluer leur éligibilité à l’inclusion et ont supprimé toutes les études inéligibles/en double. Tout désaccord concernant l’inclusion d’un article a été résolu par discussion jusqu’à ce qu’un accord soit trouvé.
Extraction et analyse des données
Français À partir des études sélectionnées, deux auteurs (NH et PAM) ont extrait indépendamment les données suivantes dans Microsoft Excel : année de publication, pays où l’étude a été menée et toutes les informations individuelles disponibles sur les cas (âge, sexe, marque du vaccin COVID-19, nombre cumulé de doses de vaccin COVID-19 administrées, jours entre la dernière administration du vaccin COVID-19 et le décès, constatations post-mortem et type de procédure post-mortem). Toute divergence dans les données a été résolue par discussion et réextraction des données. Étant donné la présence de certaines données manquantes, toutes les informations disponibles ont été utilisées pour calculer des statistiques descriptives. L’âge estimé (âge exact non indiqué) et le temps déduit entre la dernière administration du vaccin et le décès (pas de temps définitif indiqué) ont été exclus des calculs et des chiffres. Étant donné que les connaissances concernant la sécurité du vaccin contre la COVID-19 ont progressé depuis les publications originales, nous avons réalisé une revue contemporaine : trois médecins experts (RH, WM, PAM) expérimentés en matière de jugement de décès et d’anatomopathologie clinique ont examiné de manière indépendante les données disponibles pour chaque cas (tableau S1), notamment les informations démographiques, la vignette clinique, les données de vaccination, les résultats d’autopsie macroscopique et histologique, et ont déterminé si la vaccination contre la COVID-19 était la cause directe ou contribuait significativement au mécanisme de décès décrit. Les médecins ont évalué les relations temporelles, la solidité des preuves et la cohérence des résultats avec caractéristiques rapportées et présentations courantes des décès associés au vaccin contre la COVID-19 documentés dans le VAERS [15], ainsi que d’autres étiologies potentielles pour statuer sur chaque cas. Un accord était atteint lorsque deux médecins ou plus jugeaient un cas de manière concordante. Pour l’étude de Chaves [23], seuls les cas liés aux systèmes cardiovasculaire et hématologique ont été jugés comme étant liés au vaccin en raison d’une forte probabilité que la vaccination contre la COVID-19 ait contribué au décès. Cette évaluation de haute probabilité a été étayée par l’évaluation de cas similaires documentés dans le VAERS, où les événements indésirables cardiovasculaires et hématologiques représentent une part importante des rapports d’événements indésirables graves [15]. Les informations sur les cas individuels manquaient pour les autres cas, limitant la décision pour les résultats non cardiovasculaires ou hématologiques.
Résultats
Une recherche dans la base de données a permis de recenser 678 études susceptibles de répondre à nos critères d’inclusion. 562 doublons ont été éliminés. Sur les 116 articles restants, 36 répondaient à nos critères d’inclusion. Une analyse plus approfondie des références nous a permis d’identifier 18 articles supplémentaires, dont 8 répondaient à nos critères d’inclusion. Au total, nous avons identifié 44 études contenant des rapports d’autopsie ou d’autopsie restreinte d’organes de personnes vaccinées contre la COVID-19 (figure 1).
Figure 1.
Diagramme de flux des éléments de rapport préférés pour les revues systématiques et les méta-analyses (PRISMA) détaillant le processus de sélection des études
Le tableau 1 résume les 44 études [19-62], qui comprennent un total de 325 cas d’autopsie et un cas d’autopsie restreinte à un organe (cœur). L’âge moyen au décès était de 70,4 ans et il y avait 139 femmes (42,6 %). La plupart ont reçu un vaccin Pfizer/BioNTech (41 %), suivi de Sinovac (37 %), AstraZeneca (13 %), Moderna (7 %), Johnson & Johnson (1 %) et Sinopharm (1 %).
Tableau 1.
Caractéristiques des études d’autopsie incluse et d’autopsie restreinte aux organes examinant les décès potentiels liés au vaccin contre la COVID-19. *Cas = Nombre de décès examinés post-mortem. **Dose = Nombre cumulé de doses de vaccin reçues. ***Période = Temps (en jours) entre l’administration du vaccin le plus récent et le décès. ~= Période déduite (Période estimée en utilisant toutes les informations disponibles, période définitive non donnée). Abréviations : MIS = Syndrome inflammatoire multisystémique ; SCD = Mort subite d’origine cardiaque ; IM = Infarctus du myocarde ; EP = Embolie pulmonaire ; AZ = AstraZeneca ; J&J = Johnson and Johnson ; VITT = Thrombocytopénie thrombotique immunitaire induite par le vaccin ; ADEM = Encéphalomyélite aiguë disséminée ; SCLS = Syndrome de fuite capillaire systémique ; SGB = Syndrome de Guillain-Barré ; MAT = Microangiopathies thrombotiques ; VI-ARDS = Syndrome de détresse respiratoire aiguë induit par le vaccin ; UIP = Pneumonie interstitielle usuelle.
Tableau 1.
suite.
Le système cardiovasculaire était le plus fréquemment atteint (49 %), suivi des systèmes hématologiques (17 %), respiratoires (11 %), multiviscéral (7 %), neurologiques (4 %), immunologiques (4 %) et gastro-intestinaux (1 %). Dans 7 % des cas, la cause du décès était soit inconnue, soit non naturelle (noyade, traumatisme crânien, etc.) ou infectieuse (figure 2). Un seul système organique était touché dans 302 cas, deux dans 3 cas, trois dans 8 cas et quatre ou plus dans 13 cas (figure 3).
Figure 2.
Proportion de cas par système organique affecté
Figure 3.
Nombre de systèmes d’organes affectés par cas
Le tableau 2 montre le nombre et la proportion de chaque cause de décès déclarée. La mort cardiaque subite était la cause de décès la plus fréquente (21,2 %), suivie de l’infarctus du myocarde (9,5 %), de l’embolie pulmonaire (9,5 %), de la cardiopathie ischémique (6,8 %), de la TTIV (5,8 %), de la pneumonie COVID-19 (5,8 %), de la myocardite/péricardite (5,5 %), de l’hémorragie cérébrale/sous-arachnoïdienne (2,8 %), de la coronaropathie (2,5 %), de l’insuffisance respiratoire (2,5 %) et de l’hémorragie inexpliquée (2,5 %).
Tableau 2.
Nombre et proportion de causes de décès signalées parmi les cas inclus
Tableau 2.
Suite
Le nombre de jours écoulés entre la vaccination et le décès était de 14,3 (moyenne), 3 (médiane) quelle que soit la dose ; 7,8 (moyenne), 3 (médiane) après une dose ; 23,2 (moyenne), 2 (médiane) après deux doses ; et 5,7 (moyenne), 2 (médiane) après trois doses. La distribution des jours écoulés entre la dernière administration du vaccin et le décès est fortement asymétrique à droite, ce qui montre que la plupart des décès sont survenus dans la semaine suivant la dernière vaccination (figure 4).
Figure 4.
Répartition du temps écoulé entre la dernière administration du vaccin et le décès parmi les cas
Français 240 décès (73,9 %) ont été jugés indépendamment par trois médecins comme étant significativement liés à la vaccination contre la COVID-19 (tableau S1). Parmi les évaluateurs, il y avait un accord indépendant complet (les trois médecins) sur le fait que la vaccination contre la COVID-19 a contribué au décès dans 203 cas (62,5 %). Le seul cas d’autopsie restreinte d’organe a été jugé comme étant lié à la vaccination avec un accord complet. Parmi les 240 décès qui ont été jugés comme étant significativement liés à la vaccination contre la COVID-19, la plupart ont reçu un vaccin Sinovac (46,3 %), suivi de Pfizer (30,1 %), AstraZeneca (14,6 %), Moderna (7,5 %), Johnson & Johnson (1,3 %), et Sinopharm (0,8 %) ; l’âge moyen du décès était de 55,8 ans ; le nombre de jours entre la vaccination et le décès était de 11,3 (moyenne), 3 (médiane) quelle que soit la dose ; et les principales causes de décès comprennent la mort cardiaque subite (35 %), l’embolie pulmonaire (12,5 %), l’infarctus du myocarde (12 %), le VITT (7,9 %), la myocardite (7,1 %), le syndrome inflammatoire multisystémique (4,6 %) et l’hémorragie cérébrale (3,8 %).
Discussion
Français Parmi tous les rapports d’autopsie publiés concernant la vaccination contre la COVID-19, nous avons constaté, par une décision indépendante, que 73,9 % des décès étaient attribuables à des syndromes mortels de lésions vaccinales contre la COVID-19 (Tableau S1). Le système cardiovasculaire était de loin le système organique le plus impliqué dans le décès, suivi des systèmes hématologique, respiratoire, multiviscéral, neurologique, immunologique et gastro-intestinal (Figure 2), avec trois systèmes organiques ou plus touchés dans 21 cas (Figure 3). La mort cardiaque subite, l’infarctus du myocarde, la myocardite, la péricardite, l’embolie pulmonaire, la TTIV, l’hémorragie cérébrale, la défaillance multiviscérale, l’insuffisance respiratoire et l’orage cytokinique étaient les causes de décès signalées dans la majorité des cas (Tableau 2). La majorité des décès sont survenus dans la semaine suivant la dernière administration du vaccin (Figure 4). Ces résultats corroborent les syndromes connus induits par le vaccin contre la COVID-19 et montrent des associations temporelles significatives entre la vaccination contre la COVID-19 et les décès impliquant plusieurs systèmes organiques, avec une implication prédominante des systèmes cardiovasculaire et hématologique. Les critères de causalité d’un point de vue épidémiologique ont été respectés, notamment la plausibilité biologique, l’association temporelle, la validité interne et externe, la cohérence, l’analogie et la reproductibilité avec chaque rapport de cas successif de décès après la vaccination contre la COVID-19 combiné à des études basées sur la population décrivant la mortalité parmi les vaccinés.
Français Nos résultats amplifient les inquiétudes concernant les effets indésirables du vaccin COVID-19 et leurs mécanismes. La myocardite induite par le vaccin COVID-19 [11,63,64] et l’infarctus du myocarde [65,66] ont été significativement bien décrits dans la littérature évaluée par des pairs, expliquant la proportion élevée de décès cardiovasculaires observés dans notre étude. Les effets délétères de la protéine Spike [5-9,14], en particulier sur le cœur [12,67], corroborent davantage ces résultats. Nos résultats mettent également en évidence l’implication de plusieurs systèmes d’organes décrite comme un syndrome inflammatoire multisystémique (MIS) signalé après la vaccination COVID-19 chez les enfants [68] et les adultes [69]. Un mécanisme possible par lequel le MIS se produit après la vaccination pourrait être la distribution systémique des LNP contenant de l’ARNm après l’administration du vaccin [14] et l’expression et la circulation systémiques de la protéine Spike qui en résultent, entraînant une inflammation à l’échelle du système. Français Une proportion significative de cas était due à des événements indésirables du système hématologique, ce qui n’est pas surprenant étant donné que la TTIV [70] et l’embolie pulmonaire (EP) [71] ont été rapportées dans la littérature comme des événements indésirables graves après la vaccination contre la COVID-19. Les décès causés par des effets indésirables sur le système respiratoire étaient également relativement fréquents dans notre revue, une constatation qui concorde avec la possibilité de développer un syndrome de détresse respiratoire aiguë (SDRA) ou une maladie pulmonaire interstitielle induite par les médicaments (DIILD) après la vaccination contre la COVID-19 [72,73]. Bien que rares parmi les cas de cette étude, des événements indésirables immunologiques [74], neurologiques [75] et gastro-intestinaux [76] peuvent toujours survenir après la vaccination contre la COVID-19 et, comme pour le système cardiovasculaire, peuvent être directement ou indirectement causés par l’expression ou la circulation systémique de la protéine Spike. Compte tenu du délai moyen (14,3 jours) de décès après la vaccination, l’association temporelle entre la vaccination contre la COVID-19 et le décès dans la plupart des cas est renforcée par la découverte que les séquences d’ARNm Spike du vaccin contre le SRAS-CoV-2 peuvent circuler dans le sang pendant au moins 28 jours après la vaccination [13]. La plupart des plateformes vaccinales déployées sont associées au décès, ce qui suggère qu’elles partagent une caractéristique commune à l’origine d’effets indésirables, qui est très probablement la protéine Spike.
Français Le grand nombre de décès induits par le vaccin contre la COVID-19 évalués dans cette revue est cohérent avec de nombreux articles qui font état d’une surmortalité après la vaccination contre la COVID-19. Pantazatos et Seligmann ont constaté que la mortalité toutes causes confondues augmentait de 0 à 5 semaines après l’injection dans la plupart des groupes d’âge, entraînant 146 000 à 187 000 décès associés au vaccin aux États-Unis entre février et août 2021 [77]. Avec des conclusions similaires, Skidmore a estimé que 278 000 personnes pourraient être décédées des suites du vaccin contre la COVID-19 aux États-Unis d’ici décembre 2021 [78]. Ces résultats inquiétants ont été davantage élucidés par Aarstad et Kvitastein, qui ont constaté que parmi 31 pays d’Europe, une plus forte couverture vaccinale contre la COVID-19 au sein de la population en 2021 était positivement corrélée à une augmentation de la mortalité toutes causes confondues au cours des neuf premiers mois de 2022 après contrôle des variables alternatives [79]. Depuis le lancement de la campagne mondiale de vaccination contre la COVID-19, une surmortalité non liée à la COVID-19 a été détectée dans de nombreux pays [80-85], suggérant une exposition indésirable fréquente au sein de la population mondiale. Pantazatos a estimé que les décès selon le VAERS sont sous-déclarés d’un facteur 20 [77]. Si l’on applique ce facteur de sous-déclaration au 5 mai ème En 2023, le rapport VAERS sur les décès liés au vaccin contre la COVID-19 s’élève à 35 324 [15], soit environ 706 480 décès aux États-Unis et dans les autres pays utilisant le VAERS. Si ce nombre de décès extrapolé était exact, cela impliquerait que les vaccins contre la COVID-19 contribuent à la surmortalité au sein des populations.
Plusieurs études ont analysé la relation de cause à effet entre les vaccins contre la COVID-19 et les décès. Maiese et al. [86] et Sessa et al. [87] ont utilisé les conclusions de leurs études incluses pour évaluer la relation de cause à effet entre la vaccination contre la COVID-19 et les décès. Dans ces études, ils ont constaté respectivement 14 et 15 décès avec un lien de cause à effet démontré avec la vaccination contre la COVID-19. Cependant, la méthodologie de collecte des conclusions utilisée dans ces études est défectueuse pour évaluer pleinement les liens de cause à effet à l’heure actuelle, en particulier avec les vaccins contre la COVID-19, car il s’agit de nouveaux produits médicaux et de nouvelles données de sécurité sont inévitables à mesure que le temps avance. Le délai moyen pour une évaluation appropriée de la sécurité et de l’efficacité d’un vaccin est d’environ 10,71 ans [88]. Par conséquent, la méthodologie de collecte des conclusions ne devrait être envisagée que pour les études publiées au moins quelques années après le développement du vaccin afin de conserver des conclusions valides. Français Par exemple, un article publié en 2021 indique que les vaccins AstraZeneca sont sûrs et n’ont aucun lien avec des événements indésirables graves, y compris la TTIV [89]. Cependant, après une période d’observation plus longue, d’autres chercheurs ont trouvé un lien entre les vaccins AstraZeneca et la TTIV mortelle [70, 90], ce qui a entraîné un retrait généralisé du marché [91]. Pomara et al. [37] ont utilisé les lignes directrices de l’Organisation mondiale de la santé sur les événements indésirables suivant la vaccination (EIPV) qui, selon nous, constituent une excellente méthode pour évaluer la causalité entre la vaccination contre la COVID-19 et le décès. Dans cet article, les chercheurs ont conclu qu’il pourrait exister un lien de causalité entre la vaccination contre la COVID-19 et le décès par TTIV. Malheureusement, nous n’avons pas pu utiliser correctement la méthodologie de l’OMS sur les EIPV et compléter la liste de contrôle requise pour nos cas inclus en raison du manque d’informations nécessaires sur les cas. Cette méthodologie nécessite des sujets décédés, des données détaillées et l’approbation du comité d’éthique de la recherche (IRB) et ne peut pas être utilisée dans une revue systématique. Tan et al. [92] ont utilisé des statistiques d’incidence pour analyser la relation entre les vaccins contre la COVID-19 et le décès, et ont constaté une incidence plus élevée d’effets secondaires graves par rapport aux essais cliniques préalables à l’homologation. Cette méthode n’est pas applicable à notre étude car les rapports d’autopsie inclus ne présentent pas de statistiques d’incidence. Étant donné que les méthodologies susmentionnées se sont avérées incompatibles avec notre étude, nous avons décidé d’utiliser des procédures d’adjudication dirigées par des professionnels de la santé ayant une expertise pertinente pour déterminer les liens possibles entre les vaccins contre la COVID-19 et le décès. Une méthodologie d’adjudication indépendante a été utilisée par Hulscher et al. [93] pour évaluer les liens de causalité entre les vaccins contre la COVID-19 et le décès, où ils ont trouvé un lien de causalité hautement probable entre la vaccination contre la COVID-19 et le décès par myocardite dans 28 cas d’autopsie.
Français De nombreuses études ont conclu que les vaccins contre la COVID-19 sont sûrs et efficaces [94-96]. Cependant, de nombreuses études ont démontré le contraire [97-102]. L’étude OpenSAFELY, qui a inclus plus d’un million d’adolescents et d’enfants, a démontré une efficacité extrêmement limitée et des dommages accrus liés à la vaccination contre la COVID-19. La myocardite et la péricardite n’ont été documentées que dans les groupes vaccinés contre la COVID-19 et non après l’infection par la COVID-19, et les accidents et les visites aux urgences, ainsi que les hospitalisations non planifiées étaient plus élevés après la première vaccination que dans les groupes non vaccinés [97]. De plus, Alessandria et al. ont constaté que les risques de décès toutes causes confondues étaient plus élevés chez les personnes vaccinées avec une et deux doses de vaccin contre la COVID-19 que chez les personnes non vaccinées et que les doses de rappel étaient inefficaces [98]. Ils ont également démontré une perte d’espérance de vie statistiquement significative chez les personnes vaccinées avec 2 ou 3/4 doses. Les sujets vaccinés avec 2 doses ont perdu 37 % de leur espérance de vie par rapport à la population non vaccinée pendant le suivi [98]. Français Les rapports publiés des essais randomisés contrôlés par placebo initiaux ont conclu que la vaccination contre la COVID-19 pourrait réduire considérablement la maladie ambulatoire de la COVID-19 [94, 95]. Cependant, Mead et al. ont souligné que les essais étaient de courte durée, empêchant l’observation d’événements indésirables potentiels après leur arrêt. Des inquiétudes post-autorisation concernant les méthodes et l’exécution des essais ont fait surface, notamment le non-respect des principes d’intention de traiter entraînant une surestimation de l’efficacité [99]. Des réanalyses ultérieures incluant des études de surveillance post-commercialisation ont indiqué des dommages importants liés à l’ARNm, comme le rapportent les documents de Pfizer révélant 1,6 million d’événements indésirables d’ici août 2022, décrivant des blessures graves et une incidence accrue de cancer [99]. À la lumière de l’émergence continue de données sur la sécurité et l’efficacité de la vaccination contre la COVID-19, des recherches supplémentaires sont essentielles. Notre étude apporte une contribution médico-légale significative à la compréhension du profil de sécurité de ces produits.
Notre étude présente toutes les limites des biais liés à la sélection des articles et à l’évaluation indépendante des cas. Trois évaluateurs indépendants ont évalué les informations relatives aux cas afin d’éviter toute influence des biais des investigateurs ou de leurs liens, le cas échéant, avec la vaccination contre la COVID-19. Les conclusions que nous avons tirées des résultats d’autopsie reposent sur l’évolution des connaissances sur les vaccins contre la COVID-19, qui diffère sensiblement de la date de publication des études référencées, rendant toute évaluation des biais pour ces études inapplicable. Nous avons inclus tous les cas, quel que soit le résultat de l’évaluation, dans les figures et les tableaux afin de synthétiser efficacement la littérature concernant toutes les études d’autopsie incluant les vaccins contre la COVID-19 comme exposition antérieure. L’omission des cas que nous avons jugés non liés à la vaccination dans nos tableaux et figures introduirait un biais de sélection important, car nous n’avons pas utilisé les conclusions des auteurs. Notre article présente toutes les limites des revues systématiques de rapports de cas déjà publiés, notamment un biais de sélection au niveau de l’orientation pour autopsie et de l’acceptation dans la littérature évaluée par les pairs. Nous pensons que les biais de publication pourraient avoir eu une influence importante sur nos résultats en raison de la campagne mondiale de vaccination de masse menée par les gouvernements, les sociétés médicales et les centres médicaux universitaires, ainsi que de la réticence des chercheurs à signaler les effets indésirables des nouveaux produits génétiques largement recommandés aux soignants et aux patients. Enfin, des variables confondantes, notamment les maladies concomitantes, les infections, les interactions médicamenteuses et d’autres facteurs non pris en compte, pourraient avoir joué un rôle dans le lien causal avec le décès.
En résumé, parmi l’ensemble des autopsies publiées réalisées après la vaccination contre la COVID-19 disponibles à ce jour, grâce à une revue contemporaine et indépendante, nous avons constaté que dans 73,9 % des cas, la vaccination contre la COVID-19 était la cause directe ou avait contribué de manière significative au décès. La cohérence observée entre les cas de cette revue et les effets indésirables graves liés aux vaccins contre la COVID-19 précédemment signalés, leurs mécanismes fatals connus, ainsi que notre décision indépendante, suggèrent qu’il existe une forte probabilité d’un lien de causalité entre les vaccins contre la COVID-19 et le décès. Les implications de notre étude s’appliquent aux cas de décès imprévus sans antécédents médicaux chez les personnes vaccinées contre la COVID-19. Nous pouvons en déduire que, dans de tels cas, le décès pourrait avoir été causé par la vaccination contre la COVID-19. Des investigations supplémentaires sont nécessaires de toute urgence pour étayer nos résultats et mieux élucider les mécanismes physiopathologiques du décès afin de stratifier le risque et d’éviter le décès pour le grand nombre de personnes ayant reçu ou recevant un ou plusieurs vaccins contre la COVID-19 à l’avenir. Des autopsies doivent être pratiquées sur toutes les personnes décédées ayant reçu un ou plusieurs vaccins contre la COVID-19. Une surveillance clinique des personnes vaccinées contre la COVID-19 est indiquée pendant au moins un an après la vaccination afin de garantir l’absence d’effets indésirables graves pouvant entraîner le décès.
Remerciements
Contributions des auteurs
Nicolas Hulscher : Conceptualisation, Investigation, Curation de données, Analyse formelle, Méthodologie, Administration de projet, Visualisation, Rédaction – brouillon original, Rédaction – révision et édition, Validation. Peter A. McCullough : Conceptualisation, Investigation, Méthodologie, Administration de projet, Supervision, Visualisation, Rédaction – ébauche originale, Rédaction – révision et édition, Validation. Roger Hodkinson et William Makis : Rédaction – révision et édition, Validation, Investigation. Richard Amerling et Paul Alexander : Rédaction – révision et édition, Validation. Heather Gessling, Harvey A. Risch et Mark Trozzi : Supervision, Visualisation, Rédaction – ébauche originale, Rédaction – révision et édition.
Conflits d’intérêts : Les Drs Alexander, Amerling, Gessling, Hodkinson, Makis, McCullough et Risch sont affiliés à The Wellness Company, à Boca Raton (Floride), et perçoivent une rémunération ou détiennent des participations au capital de cette société. Ces personnes n’ont joué aucun rôle dans le financement, l’analyse ou la publication de ces travaux. Aucune déclaration n’est requise pour le Dr Trozzi et M. Hulscher.
Déclaration de consentement éclairé : Non applicable car il s’agit d’un article de synthèse contenant des recherches accessibles au public.
Financement : Aucun.
Déclaration de disponibilité des données : Toutes les données extraites et analysées des études incluses sont accessibles au public.
Références
1
Tableau de bord de l’OMS sur le coronavirus (COVID-19) [Internet]. Organisation mondiale de la Santé ; [consulté le 17 mai 2023]. Disponible sur : https://covid19.who.int/
2
Lyons-Weiler J. L’utilisation de valeurs CT arbitraires et généralisées dans les tests de PCR non quantitative par transcriptase inverse (RT-PCR non quantitative) pour la COVID-19 doit cesser. Science, politique de santé publique et droit. 1er novembre 2022 ; v4.2019-2024
3
Kuter BJ, Offit PA, Poland GA. Développement de vaccins contre la COVID-19 aux États-Unis : pourquoi et comment si rapidement ? Vaccine. 28 avril 2021 ; 39(18) : 2491-2495. doi : 10.1016/j.vaccine.2021.03.077. Publication en ligne du 26 mars 2021. PMID : 33824043 ; PMCID : PMC7997594 https://doi.org/10.1016/j.vaccine.2021.03.077
4
Français Graña C, Ghosn L, Evrenoglou T, Jarde A, Minozzi S, Bergman H, Buckley BS, Probyn K, Villanueva G, Henschke N, Bonnet H, Assi R, Menon S, Marti M, Devane D, Mallon P, Lelievre JD, Askie LM, Kredo T, Ferrand G, Davidson M, Riveros C, Tovey D, Meerpohl JJ, Grasselli G, Rada G, Hróbjartsson A, Ravaud P, Chaimani A, Boutron I. Efficacité et sécurité des vaccins contre la COVID-19. Cochrane Database Syst Rev. 7 décembre 2022 ; 12(12) : CD015477. doi : 10.1002/14651858.CD015477. PMID : 36473651 ; PMCID : PMC9726273. https://doi.org/10.1002/14651858.CD015477
5
Trougakos IP, Terpos E, Alexopoulos H, Politou M, Paraskevis D, Scorilas A, Kastritis E, Andreakos E, Dimopoulos MA. Effets indésirables des vaccins à ARNm contre la COVID-19 : l’hypothèse du spike. Tendances Mol Med. 2022 juil;28(7):542-554. doi: 10.1016/j.molmed.2022.04.007. Epub 2022 21 avril. PMID : 35537987 ; ID PMC : PMC9021367. https://doi.org/10.1016/j.molmed.2022.04.007
6
Seneff S, Nigh G, Kyriakopoulos AM, McCullough PA. Suppression immunitaire innée par les vaccins à ARNm contre le SARS-CoV-2 : rôle des G-quadruplexes, des exosomes et des microARN. Food Chem Toxicol. Juin 2022 ; 164 : 113008. doi : 10.1016/j.fct.2022.113008. Publication en ligne du 15 avril 2022. PMID : 35436552 ; PMCID : PMC9012513. https://doi.org/10.1016/j.fct.2022.113008
7
Uversky VN, Redwan EM, Makis W, Rubio-Casillas A. Les anticorps IgG4 induits par des vaccinations répétées peuvent générer une tolérance immunitaire à la protéine Spike du SARS-CoV-2. Vaccines (Bâle). 17 mai 2023 ; 11(5) : 991. doi : 10.3390/vaccines11050991. PMID : 37243095 ; PMCID : PMC10222767. https://doi.org/10.3390/vaccines11050991
8
Theoharides TC. La protéine Spike du SARS-CoV-2 pourrait-elle être responsable du syndrome de la COVID longue ? Mol Neurobiol. Mars 2022 ; 59(3) : 1850-1861. doi : 10.1007/s12035-021-02696-0. Publication en ligne du 13 janvier 2022. PMID : 35028901 ; PMCID : PMC8757925. https://doi.org/10.1007/s12035-021-02696-0
9
Theoharides TC, Conti P. Attention à la protéine Spike du SARS-CoV-2 : il y a plus qu’il n’y paraît. J Biol Regul Homeost Agents. Mai-juin 2021 ; 35(3) : 833-838. doi : 10.23812/THEO_EDIT_3_21. PMID : 34100279. https://doi.org/10.23812/THEO_EDIT_3_21
10
Aleem A, Nadeem AJ. Thrombopénie thrombotique immunitaire induite par le vaccin contre le coronavirus (COVID-19). 3 octobre 2022. Dans : StatPearls [Internet]. Treasure Island (Floride) : StatPearls Publishing ; janv. 2023. PMID : 34033367.
11
Bozkurt B, Kamat I, Hotez PJ. Myocardite avec les vaccins à ARNm contre la COVID-19. Circulation. 10 août 2021 ; 144(6) : 471-484. doi : 10.1161/CIRCULATIONAHA.121.056135. Publication en ligne du 20 juillet 2021. PMID : 34281357 ; PMCID : PMC8340726. https://doi.org/10.1161/CIRCULATIONAHA.121.056135
12
Yonker LM, Swank Z, Bartsch YC, Burns MD, Kane A, Boribong BP, Davis JP, Loiselle M, Novak T, Senussi Y, Cheng CA, Burgess E, Edlow AG, Chou J, Dionne A, Balaguru D, Lahoud-Rahme M, Arditi M, Julg B, Randolph AG, Alter G, Fasano A, Walt DR. Protéine de spicule circulante détectée dans la myocardite post-vaccinale à ARNm COVID-19. Circulation. 14 mars 2023 ; 147(11) : 867-876. doi : 10.1161/CIRCULATIONAHA.122.061025. Publication en ligne du 4 janvier 2023. PMID : 36597886 ; PMCID : PMC10010667. https://doi.org/10.1161/CIRCULATIONAHA.122.061025
13
Castruita JAS, Schneider UV, Mollerup S, Leineweber TD, Weis N, Bukh J, Pedersen MS, Westh H. Les séquences d’ARNm de la protéine Spike du vaccin contre le SARS-CoV-2 circulent dans le sang jusqu’à 28 jours après la vaccination contre la COVID-19. APMIS. Mars 2023 ; 131(3) : 128-132. doi : 10.1111/apm.13294. Publication en ligne du 29 janvier 2023. PMID : 36647776 ; PMCID : PMC10107710. https://doi.org/10.1111/apm.13294
14
Parry PI, Lefringhausen A, Turni C, et al. « Spikeopathie » : la protéine Spike de la COVID-19 est pathogène, d’après l’ARNm du virus et du vaccin. Biomedicines. 2023 ; 11(8) : 2287. Publié le 17 août 2023. doi:10.3390/biomedicines11082287 https://doi.org/10.3390/biomedicines11082287
15
Système de déclaration des effets indésirables des vaccins (VAERS) [en ligne]. Disponible à l’adresse : https://vaers.hhs.gov .
16
Harald Walach, Rainer J. Klement et Wouter Aukema. 2021. La sécurité des vaccins contre la COVID-19 : faut-il repenser la politique ? Sci, Publ Health Pol & Law 3:87-99.
17
Scarl R, Parkinson B, Arole V, Hardy T, Allenby P. L’autopsie à l’hôpital : l’importance de maintenir l’autopsie comme option. Autopsy Case Rep. 17 févr. 2022 ; 12 : e2021333. doi : 10.4322/acr.2021.333. PMID : 35252044 ; PMCID : PMC8890781. https://doi.org/10.4322/acr.2021.333
18
Moher D, Liberati A, Tetzlaff J, Altman DG ; Groupe PRISMA. Éléments de rapport privilégiés pour les revues systématiques et les méta-analyses : la déclaration PRISMA. J Clin Epidemiol. Oct. 2009 ; 62(10) : 1006-12. doi : 10.1016/j.jclinepi.2009.06.005. Publication en ligne du 23 juillet 2009. PMID : 19631508. https://doi.org/10.1016/j.jclinepi.2009.06.005
19
Hojberg Y, Abdeljaber M, Prahlow JA. Éosinophilie généralisée après administration du vaccin Moderna contre la COVID-19 : rapport de cas. Acad Forensic Pathol. 2023 mars ; 13(1): 9-15. doi : 10.1177/19253621231157933. Publication en ligne du 28 mars 2023. PMID : 37091194 ; PMCID : PMC10119868. https://doi.org/10.1177/19253621231157933
20
Nushida H, Ito A, Kurata H, Umemoto H, Tokunaga I, Iseki H, Nishimura A. Un cas d’inflammation multi-organique mortelle suite à la vaccination contre la COVID-19. Jambe avec (Tokyo). 20 mars 2023 ;63 :102244. doi: 10.1016/j.legalmed.2023.102244. Publication électronique avant impression. PMID : 36990036 ; ID PMC : PMC10027302. https://doi.org/10.1016/j.legalmed.2023.102244
21
Jeon YH, Choi S, Park JH, Lee JK, Yeo NS, Lee S, Suh YL. Mort subite associée à de possibles poussées de sclérose en plaques après vaccination et infection par la COVID-19 : rapport de cas et revue de la littérature. J Korean Med Sci. 13 mars 2023 ; 38(10) : e78. doi : 10.3346/jkms.2023.38.e78. PMID : 36918031 ; PMCID : PMC10010908. https://doi.org/10.3346/jkms.2023.38.e78
22
Esposito M, Cocimano G, Vanaria F, Sessa F, Salerno M. Décès dû à la COVID-19 chez un sujet entièrement vacciné : rapport d’autopsie complet. Vaccines (Bâle). 9 janvier 2023 ; 11(1):142. doi : 10.3390/vaccines11010142. PMID : 36679987 ; PMCID : PMC9865400. https://doi.org/10.3390/vaccines11010142
23
Chaves JJ, Bonilla JC, Chaves-Cabezas V, Castro A, Polo JF, Mendoza O, Correa-Rodríguez J, Piedrahita AC, Romero-Fandiño IA, Caro MV, González AC, Sánchez LK, Murcia F, Márquez G, Benavides A, Quiroga MDP, López J, Parra-Medina R. Une étude post-mortem de patients vaccinés contre le SRAS-CoV-2 en Colombie. Rév. Esp Patol. 2023 janv.-mars;56(1):4-9. doi: 10.1016/j.patol.2022.09.003. Publication électronique du 31 octobre 2022. PMID : 36599599 ; ID PMC : PMC9618417. https://doi.org/10.1016/j.patol.2022.09.003
24
Mörz M. Rapport de cas : Encéphalite nécrosante multifocale et myocardite après vaccination par l’ARNm BNT162b2 contre la COVID-19. Vaccines (Bâle). 1er octobre 2022 ; 10(10) : 1651. doi : 10.3390/vaccines10101651. PMID : 36298516 ; PMCID : PMC9611676. https://doi.org/10.3390/vaccines10101651
25
Alunni V, Bernardi C, Chevalier N, Cabusat C, Quatrehomme G, Torrents J, Biglia E, Gaillard Y, Drici MD. Des anticorps anti-PF4 post-mortem confirment un cas rare de syndrome de thrombose thrombocytopénie associé à la vaccination anti-COVID-19 ChAdOx1. Int J Legal Med. 2023 mars ; 137(2) : 487-492. doi : 10.1007/s00414-022-02910-1. Publication en ligne du 27 octobre 2022. PMID : 36289074 ; PMCID : PMC9607767. https://doi.org/10.1007/s00414-022-02910-1
26
Takahashi M, Kondo T, Yamasaki G, Sugimoto M, Asano M, Ueno Y, Nagasaki Y. Rapport d’autopsie d’une dissection aortique compliquée d’une péricardite histiolymphocytaire et d’une inflammation aortique après une vaccination à ARNm contre la COVID-19. Leg Med (Tokyo). 2022 novembre ;59 :102154. doi: 10.1016/j.legalmed.2022.102154. Epub 29 septembre 2022. PMID : 36191411 ; ID PMC : PMC9519380. https://doi.org/10.1016/j.legalmed.2022.102154
27
Murata K, Nakao N, Ishiuchi N, Fukui T, Katsuya N, Fukumoto W, Oka H, Yoshikawa N, Nagao T, Namera A, Kakimoto N, Oue N, Awai K, Yoshimoto K, Nagao M. Quatre cas de choc cytokinique après vaccination contre la COVID-19 : rapport de cas. Front Immunol. 15 août 2022 ; 13:967226. doi : 10.3389/fimmu.2022.967226. PMID : 36045681 ; PMCID : PMC9420842. https://doi.org/10.3389/fimmu.2022.967226
28
Satomi H, Katano H, Kanno H, Kobayashi M, Ohkuma Y, Hashidume N, Usui T, Tsukada S, Ito I. Un cas d’autopsie de myocardite fulminante après inoculation du vaccin contre le coronavirus 2 du syndrome respiratoire aigu sévère. Pathol Int. 2022 oct;72(10):519-524. doi: 10.1111/pin.13267. Epub 30 août 2022. PMID : 36040128 ; ID PMC : PMC9537995. https://doi.org/10.1111/pin.13267
29
Suzuki H, Ro A, Takada A, Saito K, Hayashi K. Résultats d’autopsie des décès post-vaccination contre la COVID-19 dans la métropole de Tokyo, Japon, 2021. Leg Med (Tokyo). 2022 nov. ; 59:102134. doi : 10.1016/j.legalmed.2022.102134. Publication en ligne du 20 août 2022. PMID : 36037554 ; PMCID : PMC9392553. https://doi.org/10.1016/j.legalmed.2022.102134
30
Mele F, Tafuri S, Stefanizzi P, D Amati A, Calvano M, Leonardelli M, Macorano E, Duma S, De Gabriele G, Introna F, De Donno A. Thrombose du sinus veineux cérébral après vaccination contre la COVID-19 et déficit congénital en facteurs de coagulation : existe-t-il une corrélation ? Hum Vaccin Immunother. 30 novembre 2022 ; 18(6) : 2095166. doi : 10.1080/21645515.2022.2095166. Publication en ligne du 27 juillet 2022. PMID : 35895937 ; PMCID : PMC9746424. https://doi.org/10.1080/21645515.2022.2095166
31
Yoshimura Y, Sasaki H, Miyata N, Miyazaki K, Okudela K, Tateishi Y, Hayashi H, Kawana-Tachikawa A, Iwashita H, Maeda K, Ihama Y, Hatayama Y, Ryo A, Tachikawa N. Un cas d’autopsie de syndrome de détresse respiratoire aiguë de type COVID-19 après vaccination contre le SARS-CoV-2 par l’ARNm-1273. Int J Infect Dis. Août 2022 ; 121 : 98-101. doi: 10.1016/j.ijid.2022.04.057. Epub 30 avril 2022. PMID : 35500794 ; ID PMC : PMC9054706. https://doi.org/10.1016/j.ijid.2022.04.057
32
Roncati L, Manenti A, Corsi L. Série de trois cas de décès thrombotiques chez des patients de plus de 50 ans présentant des comorbidités temporaires après la vaccination contre la COVID-19 par modARN. Pathogens. 3 avril 2022 ; 11(4) : 435. doi : 10.3390/pathogens11040435. PMID : 35456110 ; PMCID : PMC9032304. https://doi.org/10.3390/pathogens11040435
33
Kang DH, Na JY, Yang JH, Moon SH, Kim SH, Jung JJ, Cha HJ, Ahn JH, Park YW, Cho SY, Yu HK, Lee SH, Park MY, Kim JW, Byun JH. Myocardite fulminante à cellules géantes après vaccination hétérologue contre ChAdOx1 nCoV-19 et Pfizer-BioNTech COVID-19. Medicina (Kaunas). 20 mars 2022 ; 58(3):449. doi : 10.3390/medicina58030449. PMID : 35334625 ; PMCID : PMC8950462. https://doi.org/10.3390/medicina58030449
34
Kamura Y, Terao T, Akao S, Kono Y, Honma K, Matsue K. Microangiopathie thrombotique mortelle avec rhabdomyolyse comme symptôme initial après la première dose du vaccin à ARNm-1273 : rapport de cas. Int J Infect Dis. 2022 avr. ; 117 : 322-325. doi : 10.1016/j.ijid.2022.02.031. Publication en ligne du 18 février 2022. PMID : 35189339 ; PMCID : PMC8853962. https://doi.org/10.1016/j.ijid.2022.02.031
35
Ishioka Y, Makiguchi T, Itoga M, Tanaka H, Taima K, Goto S, Tasaka S. Exacerbation aiguë d’une pneumopathie interstitielle après vaccination contre le SRAS-CoV-2 : une série de cas. Chest. Déc. 2022 ; 162(6) : e311-e316. doi : 10.1016/j.chest.2022.08.2213. PMID : 36494131 ; PMCID : PMC9723271. https://doi.org/10.1016/j.chest.2022.08.2213
36
Gill JR, Tashjian R, Duncanson E. Constatations histopathologiques cardiaques d’autopsie chez deux adolescents après la deuxième dose du vaccin contre la COVID-19. Arch Pathol Lab Med. 1er août 2022 ; 146(8) : 925-929. doi : 10.5858/arpa.2021-0435-SA. PMID : 35157759. https://doi.org/10.5858/arpa.2021-0435-SA
37
Pomara C, Salerno M, Esposito M, Sessa F, Certo F, Tripodo C, Rappa F, Barbagallo GM. Résultats histologiques et immunohistochimiques dans un cas mortel de thrombocytopénie thrombotique après vaccination contre ChAdOx1 nCov-19. Pathol Res Pract. 2022 mars ; 231 : 153796. doi : 10.1016/j.prp.2022.153796. Publication en ligne du 4 février 2022. PMID : 35144085. https://doi.org/10.1016/j.prp.2022.153796
38
Yeo A, Kuek B, Lau M, Tan SR, Chan S. Décès post-vaccinaux contre la COVID-19 – Expérience précoce de Singapour. Forensic Sci Int. 19 janvier 2022 ; 332 : 111199. doi : 10.1016/j.forsciint.2022.111199. Publication électronique avant impression. PMID : 35078041 ; PMCID : PMC8767909. https://doi.org/10.1016/j.forsciint.2022.111199
39
Ameratunga R, Woon ST, Sheppard MN, Garland J, Ondruschka B, Wong CX, Stewart RAH, Tatley M, Stables SR, Tse RD. Premier cas identifié de myocardite nécrosante éosinophile fulminante mortelle après l’administration de la première dose du vaccin à ARNm Pfizer-BioNTech contre la COVID-19 (BNT162b2, Comirnaty) : une réaction d’hypersensibilité idiosyncrasique extrêmement rare. J Clin Immunol. 2022 avr. ; 42(3) : 441-447. doi : 10.1007/s10875-021-01187-0. Publication en ligne du 3 janvier 2022. PMID : 34978002 ; PMCID : PMC8720536. https://doi.org/10.1007/s10875-021-01187-0
40
Günther A, Brämer D, Pletz MW, Kamradt T, Baumgart S, Mayer TE, Baier M, Autsch A, Mawrin C, Schönborn L, Greinacher A, Thiele T. Rapport de cas de thrombopénie immunitaire thrombotique compliquée à long terme induite par un vaccin (A). Vaccines (Bâle). 17 novembre 2021 ; 9(11) : 1344. doi : 10.3390/vaccines9111344. PMID : 34835275 ; PMCID : PMC8622649. https://doi.org/10.3390/vaccines9111344
41
Permezel F, Borojevic B, Lau S, de Boer HH. Encéphalomyélite aiguë disséminée (EMAD) suite à une récente vaccination contre la COVID-19 par Oxford/AstraZeneca. Forensic Sci Med Pathol. Mars 2022 ; 18(1) : 74-79. doi : 10.1007/s12024-021-00440-7. Publication en ligne le 4 novembre 2021. PMID : 34735684 ; PMCID : PMC8567127. https://doi.org/10.1007/s12024-021-00440-7
42
Choi S, Lee S, Seo JW, Kim MJ, Jeon YH, Park JH, Lee JK, Yeo NS. Mort subite induite par myocardite après vaccination contre la COVID-19 par l’ARNm BNT162b2 en Corée : étude de cas axée sur les résultats histopathologiques. J Korean Med Sci. 18 octobre 2021 ; 36(40) : e286. doi : 10.3346/jkms.2021.36.e286. PMID : 34664804 ; PMCID : PMC8524235. https://doi.org/10.3346/jkms.2021.36.e286
43
Schneider J, Sottmann L, Greinacher A, Hagen M, Kasper HU, Kuhnen C, Schlepper S, Schmidt S, Schulz R, Thiele T, Thomas C, Schmeling A. Enquête post-mortem sur les décès consécutifs à la vaccination contre la COVID-19. Int J Legal Med. 2021 Nov;135(6):2335-2345. doi: 10.1007/s00414-021-02706-9. Publication en ligne du 30 sept. 2021. PMID: 34591186; PMCID: PMC8482743. https://doi.org/10.1007/s00414-021-02706-9
44
Schneider J, Sottmann L, Greinacher A, Hagen M, Kasper HU, Kuhnen C, Schlepper S, Schmidt S, Schulz R, Thiele T, Thomas C, Schmeling A. Enquête post-mortem sur les décès consécutifs à la vaccination contre la COVID-19. Int J Legal Med. 2021 Nov;135(6):2335-2345. doi: 10.1007/s00414-021-02706-9. Publication en ligne du 30 sept. 2021. PMID: 34591186; PMCID: PMC8482743. https://doi.org/10.1056/NEJMc2109975
45
Wiedmann M, Skattør T, Stray-Pedersen A, Romundstad L, Antal EA, Marthinsen PB, Sørvoll IH, Leiknes Ernstsen S, Lund CG, Holme PA, Johansen TO, Brunborg C, Aamodt AH, Schultz NH, Skagen K, Skjelland M. Thrombocytopénie thrombotique immunitaire induite par un vaccin provoquant une forme grave de thrombose veineuse cérébrale avec un taux de mortalité élevé : une série de cas. Neurol. Frontal. 30 juillet 2021 ; 12 : 721146. doi: 10.3389/fneur.2021.721146. PMID : 34393988 ; ID PMC : PMC8363077. https://doi.org/10.3389/fneur.2021.721146
46
Pomara C, Sessa F, Ciaccio M, Dieli F, Esposito M, Giammanco GM, Garozzo SF, Giarratano A, Prati D, Rappa F, Salerno M, Tripodo C, Mannucci PM, Zamboni P. Vaccin contre la COVID-19 et décès : algorithme de causalité selon le diagnostic d’éligibilité de l’OMS. Diagnostic (Bâle). 26 mai 2021 ;11(6):955. doi: 10.3390/diagnostics11060955. PMID : 34073536 ; ID PMC : PMC8229116. https://doi.org/10.3390/diagnostics11060955
47
Althaus K, Möller P, Uzun G, Singh A, Beck A, Bettag M, Bösmüller H, Guthoff M, Dorn F, Petzold GC, Henkes H, Heyne N, Jumaa H, Kreiser K, Limpach C, Luz B, Maschke M, Müller JA, Münch J, Nagel S, Pötzsch B, Müller J, Schlegel C, Viardot A, Bäzner H, Wolf M, Pelzl L, Warm V, Willinek WA, Steiner J, Schneiderhan-Marra N, Vollherbst D, Sachs UJ, Fend F, Bakchoul T. Plaquettes procoagulantes à médiation par anticorps dans la thrombocytopénie thrombotique immunitaire associée à la vaccination contre le SRAS-CoV-2. Hématologique. 1er août 2021 ; 106(8) : 2170-2179. doi: 10.3324/haematol.2021.279000. PMID : 34011137 ; ID PMC : PMC8327736. https://doi.org/10.3324/haematol.2021.279000
48
Edler C, Klein A, Schröder AS, Sperhake JP, Ondruschka B. Décès associés au nouveau vaccin contre le SARS-CoV-2 (Comirnaty®). Leg Med (Tokyo). Juil. 2021 ; 51:101895. doi : 10.1016/j.legalmed.2021.101895. Publication en ligne du 17 avril 2021. PMID : 33895650 ; PMCID : PMC8052499. https://doi.org/10.1016/j.legalmed.2021.101895
49
Hansen T, Titze U, Kulamadayil-Heidenreich NSA, Glombitza S, Tebbe JJ, Röcken C, Schulz B, Weise M, Wilkens L. Premier cas d’étude post-mortem chez un patient vacciné contre le SARS-CoV-2. Int J Infect Dis. Juin 2021 ; 107 : 172-175. doi : 10.1016/j.ijid.2021.04.053. Publication en ligne du 16 avril 2021. PMID : 33872783 ; PMCID : PMC8051011. https://doi.org/10.1016/j.ijid.2021.04.053
50
Baronti A, Gentile F, Manetti AC, Scatena A, Pellegrini S, Pucci A, Franzini M, Castiglione V, Maiese A, Giannoni A, Pistello M, Emdin M, Aquaro GD, Di Paolo M. Myocardial Infarction Following COVID-19 Vaccine Administration: Post Hoc, Ergo Propter Hoc? Viruses. 2022 Jul 27;14(8):1644. doi: 10.3390/v14081644. PMID: 36016266; PMCID: PMC9413746. https://doi.org/10.3390/v14081644
51
Ittiwut C, Mahasirimongkol S, Srisont S, Ittiwut R, Chockjamsai M, Durongkadech P, Sawaengdee W, Khunphon A, Larpadisorn K, Wattanapokayakit S, Wetchaphanphesat S, Arunotong S, Srimahachota S, Pittayawonganon C, Thammawijaya P, Sutdan D, Doungngern P, Khongphatthanayothin A, Kerr SJ, Shotelersuk V. Genetic basis of sudden death after COVID-19 vaccination in Thailand. Heart Rhythm. 2022 Aug 5;19(11):1874–9. doi: 10.1016/j.hrthm.2022.07.019. Epub ahead of print. PMID: 35934244; PMCID: PMC9352648. https://doi.org/10.1016/j.hrthm.2022.07.019
52
Greinacher A, Thiele T, Warkentin TE, Weisser K, Kyrle PA, Eichinger S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N Engl J Med. 2021 Jun 3;384(22):2092-2101. doi: 10.1056/NEJMoa2104840. Epub 2021 Apr 9. PMID: 33835769; PMCID: PMC8095372. https://doi.org/10.1056/NEJMoa2104840
53
Mauriello A, Scimeca M, Amelio I, Massoud R, Novelli A, Di Lorenzo F, Finocchiaro S, Cimino C, Telesca R, Chiocchi M, Sun Q, Wang Y, Shi Y, Novelli G, Melino G. Thromboembolie après vaccination contre la COVID-19 chez les patients présentant une thrombocytopénie préexistante. Maladie de la mort cellulaire 3 août 2021 ;12(8):762. doi: 10.1038/s41419-021-04058-z. PMID : 34344867 ; ID PMC : PMC8328816. https://doi.org/10.1038/s41419-021-04058-z
54
Bjørnstad-Tuveng TH, Rudjord A, Anker P. Hémorragie cérébrale mortelle après le vaccin contre la COVID-19. Journal de l’Association médicale norvégienne. 29 avril 2021 ; 141. Anglais, norvégien. doi: 10.4045/tidsskr.21.0312. PMID : 33928772. https://doi.org/10.4045/tidsskr.21.0312
55
Scully M, Singh D, Lown R, Poles A, Solomon T, Levi M, Goldblatt D, Kotoucek P, Thomas W, Lester W. Anticorps pathologiques dirigés contre le facteur plaquettaire 4 après vaccination contre le ChAdOx1 nCoV-19. N Engl J Med. 10 juin 2021 ; 384(23) : 2202-2211. doi : 10.1056/NEJMoa2105385. Publication en ligne du 16 avril 2021. PMID : 33861525 ; PMCID : PMC8112532. https://doi.org/10.1056/NEJMoa2105385
56
Choi GJ, Baek SH, Kim J, Kim JH, Kwon GY, Kim DK, Jung YH, Kim S. Syndrome de fuite capillaire systémique fatal après vaccination contre le SARS-CoV-2 chez un patient atteint de myélome multiple. Emerg Infect Dis. 2021 Nov;27(11):2973-2975. doi: 10.3201/eid2711.211723. Epub 30 août 2021. PMID: 34459725; PMCID: PMC8544977. https://doi.org/10.3201/eid2711.211723
57
Schwab C, Domke LM, Hartmann L, Stenzinger A, Longerich T, Schirmacher P. Caractérisation histopathologique autopsiée de la myocardite après vaccination anti-SARS-CoV-2. Clin Res Cardiol. Mars 2023 ; 112(3) : 431-440. doi : 10.1007/s00392-022-02129-5. Publication en ligne du 27 novembre 2022. PMID : 36436002 ; PMCID : PMC9702955. https://doi.org/10.1007/s00392-022-02129-5
58
Hirschbühl K, Schaller T, Märkl B, Claus R, Sipos E, Rentschler L, Maccagno A, Grosser B, Kling E, Neidig M, Kröncke T, Spring O, Braun G, Bösmüller H, Seidl M, Esposito I, Pablik J, Hilsenbeck J, Boor P, Beer M, Dintner S, Wylezich C. Charges virales élevées : qu’est-ce qui entraîne des cas mortels de COVID-19 chez les personnes vaccinées ? – une étude d’autopsie. Mod Pathol. Août 2022 ; 35(8) : 1013-1021. doi : 10.1038/s41379-022-01069-9. Publication en ligne du 1er avril 2022. PMID : 35365771 ; PMCID : PMC8974809. https://doi.org/10.1038/s41379-022-01069-9
59
Hoshino N, Yanase M, Ichiyasu T, Kuwahara K, Kawai H, Muramatsu T, Ishii H, Tsukamoto T, Morimoto SI, Izawa H. An autopsy case report of fulminant myocarditis: Following mRNA COVID-19 vaccination. J Cardiol Cases. 2022 Dec;26(6):391-394. doi: 10.1016/j.jccase.2022.06.006. Epub 2022 Jul 4. PMID: 35812802; PMCID: PMC9250935. https://doi.org/10.1016/j.jccase.2022.06.006
60
Colombo D, Del Nonno F, Marchioni L, Lalle E, Gallì P, Vaia F, Falasca L. Autopsies Revealed Pathological Features of COVID-19 in Unvaccinated vs. Vaccinated Patients. Biomedicines. 2023 Feb 14;11(2):551. doi: 10.3390/biomedicines11020551. PMID: 36831087; PMCID: PMC9953314. https://doi.org/10.3390/biomedicines11020551
61
Mosna K, Vadkerti P, Papp L, Palkovic M, Janega P, Babal P. Syndrome de Guillain-Barré avec issue fatale après vaccination contre la Covid-19 – rapport de cas confirmé par autopsie. The Open Neurology Journal. 10 mars 2022 ; 16(1). doi:10.2174/1874205x-v16-e2207270 https://doi.org/10.2174/1874205x-v16-e2207270
62
Kaimori R, Nishida H, Uchida T, Tamura M, Kuroki K, Murata K, Hatakeyama K, Ikeda Y, Amemiya K, Nishizono A, Daa T, Mori S. Distribution histopathologique de type TMA des thromboses multiviscérales après l’administration initiale du vaccin à ARNm BNT162b2 (Comirnaty, Pfizer/BioNTech) : rapport d’autopsie. Thromb J. 6 oct. 2022 ; 20(1): 61. doi : 10.1186/s12959-022-00418-7. PMID : 36203145 ; PMCID : PMC9540301. https://doi.org/10.1186/s12959-022-00418-7
63
Français Wong HL, Hu M, Zhou CK, Lloyd PC, Amend KL, Beachler DC, Secora A, McMahill-Walraven CN, Lu Y, Wu Y, Ogilvie RP, Reich C, Djibo DA, Wan Z, Seeger JD, Akhtar S, Jiao Y, Chillarige Y, Do R, Hornberger J, Obidi J, Forshee R, Shoaibi A, Anderson SA. Risque de myocardite et de péricardite après la vaccination à ARNm contre la COVID-19 aux États-Unis : une étude de cohorte dans les bases de données de demandes d’indemnisation. Lancet. 11 juin 2022 ; 399(10342) : 2191-2199. doi : 10.1016/S0140-6736(22)00791-7. PMID : 35691322 ; PMCID : PMC9183215. https://doi.org/10.1016/S0140-6736(22)00791-7
64
Park DY, An S, Kaur A, Malhotra S, Vij A. Myocardite après vaccination à ARNm contre la COVID-19 : revue systématique des rapports de cas et des séries de cas. Clin Cardiol. Juil. 2022 ; 45(7) : 691-700. doi : 10.1002/clc.23828. Publication en ligne du 2 juin 2022. PMID : 35652390 ; PMCID : PMC9286338. https://doi.org/10.1002/clc.23828
65
Baqi DH, Kakamad FH, Mahmood ZH, Fattah FH, Ahmed SF, Hassan MN, Hama Amin BJ, Mohammed SH, Mikael TM, Hassan HA, Salh AM. Infarctus du myocarde suite à l’administration du vaccin contre la COVID-19 ; une revue systématique. Hélion. 11 novembre 2022 ; 8(11) : e11385. doi: 10.1016/j.helion.2022.e11385. PMID : 36406668 ; ID PMC : PMC9650518. https://doi.org/10.1016/j.helion.2022.e11385
66
Zafar U, Zafar H, Ahmed MS, Khattak M. Lien entre les vaccins contre la COVID-19 et l’infarctus du myocarde. World J Clin Cases. 6 octobre 2022 ; 10(28) : 10109-10119. doi : 10.12998/wjcc.v10.i28.10109. PMID : 36246837 ; PMCID : PMC9561578. https://doi.org/10.12998/wjcc.v10.i28.10109
67
Français Baumeier C, Aleshcheva G, Harms D, Gross U, Hamm C, Assmus B, Westenfeld R, Kelm M, Rammos S, Wenzel P, Munzel T, Elsässer A, Gailani M, Perings C, Bourakkadi A, Flesch M, Kempf T, Bauersachs J, Escher F, Schultheiss HP. Inflammation intramyocardique après vaccination contre la COVID-19 : une série de cas prouvés par biopsie endomyocardique. Int J Mol Sci. 22 juin 2022 ; 23(13):6940. doi: 10.3390/ijms23136940. PMID : 35805941 ; ID PMC : PMC9266869. https://doi.org/10.3390/ijms2
68
Wangu Z, Swartz H, Doherty M. Syndrome inflammatoire multisystémique chez l’enfant (MIS-C) possiblement secondaire à la vaccination à ARNm contre la COVID-19. BMJ Case Rep. 30 mars 2022 ; 15(3) : e247176. doi : 10.1136/bcr-2021-247176. PMID : 35354564 ; PMCID : PMC8968554. https://doi.org/10.1136/bcr-2021-247176
69
Ehikhametalor K, Deans-Minott J, Duncan JP. Syndrome inflammatoire multisystémique de l’adulte (MIS-A) après infection par la COVID-19 et vaccination récente par un vecteur adénoviral recombinant codant pour l’antigène de la protéine de spicule du SARS-CoV-2 (ChAdOx1 nCoV-19, Vaxzevria). J Intensive Care Med. Févr. 2023 ; 38(2) : 232-237. doi : 10.1177/08850666221121589. Publication en ligne du 17 août 2022. PMID : 35979616 ; PMCID : PMC9389272. https://doi.org/10.1177/08850666221121589
70
Zidan A, Noureldin A, Kumar SA, Elsebaie A, Othman M. Thrombose immunitaire et thrombopénie associées au vaccin contre la COVID-19 (TTIV) : divergences diagnostiques et implications mondiales. Semin Thromb Hemost. Fév. 2023 ; 49(1) : 9-14. doi : 10.1055/s-0042-1759684. Publication en ligne le 5 janvier 2023. PMID : 36603593. https://doi.org/10.1055/s-0042-1759684
71
Ifeanyi N, Chinenye N, Oladiran O, David E, Mmonu C, Ogbonna-Nwosu C. Embolie pulmonaire isolée après vaccination contre la COVID-19 : deux rapports de cas et revue des complications et du suivi de l’embolie pulmonaire post-aiguë. J Community Hosp Intern Med Perspect. 15 novembre 2021 ; 11(6) : 877-879. doi : 10.1080/20009666.2021.1990825. PMID : 34804412 ; PMCID : PMC8604520. https://doi.org/10.1080/20009666.2021.1990825
72
Abraham B, Mohammed Saeed H, Azeez Pasha SA. Syndrome de détresse respiratoire aiguë secondaire à l’administration du vaccin à ARNm contre la COVID-19 chez une femme enceinte : rapport de cas. Qatar Med J. 9 août 2022 ; 2022(3):40. doi : 10.5339/qmj.2022.40. PMID : 35974885 ; PMCID : PMC9372495. https://doi.org/10.5339/qmj.2022.40
73
Yoshifuji A, Ishioka K, Masuzawa Y, Suda S, Murata S, Uwamino Y, Fujino M, Miyahara H, Hasegawa N, Ryuzaki M, Hoshino H, Sekine K. Maladie pulmonaire interstitielle induite par le vaccin contre la COVID-19. J Infect Chemother. 2022 janv.;28(1):95-98. doi: 10.1016/j.jiac.2021.09.010. Epub 2021 20 sept. PMID : 34580010 ; ID PMC : PMC8450284. https://doi.org/10.1016/j.jiac.2021.09.010
74
Chen Y, Xu Z, Wang P, Li XM, Shuai ZW, Ye DQ, Pan HF. Phénomènes auto-immuns d’apparition récente après la vaccination contre la COVID-19. Immunologie. 2022 avr. ; 165(4) : 386-401. doi : 10.1111/imm.13443. Publication en ligne le 7 janvier 2022. PMID : 34957554. https://doi.org/10.1111/imm.13443
75
Hosseini R, Askari N. Revue des effets secondaires neurologiques de la vaccination contre la COVID-19. Eur J Med Res. 25 févr. 2023 ; 28(1):102. doi : 10.1186/s40001-023-00992-0. PMID : 36841774 ; PMCID : PMC9959958. https://doi.org/10.1186/s40001-023-00992-0
76
Ajmera K, Bansal R, Wilkinson H, Goyal L. Complications gastro-intestinales des vaccins contre la COVID-19. Cureus. 12 avril 2022 ; 14(4) : e24070. doi : 10.7759/cureus.24070. PMID : 35573556 ; PMCID : PMC9097558. https://doi.org/10.7759/cureus.24070
77
Pantazatos S, Seligmann H. Vaccination contre la COVID-19 et risque de mortalité toutes causes confondues stratifié par âge. Research Gate, 26 octobre 2021. Publication en ligne le 26 octobre. DOI : 10.13140/RG.2.2.28257.43366. https://doi.org/10.13140/RG.2.2.28257.43366
78
Skidmore, M. 2023. Les expériences de la maladie et de la vaccination contre la COVID-19 dans les cercles sociaux influencent les décisions de vaccination contre la COVID-19. Science, politique de santé publique et droit 4 : 208-226.
79
Aarstad, J.; Kvitastein, OA. Existe-t-il un lien entre le taux de vaccination contre la COVID-19 en Europe en 2021 et la surmortalité toutes causes confondues en 2022 ? Preprints.org 2023, 2023020350. doi:10.20944/preprints202302.0350.v1. https://doi.org/10.20944/preprints202302.0350.v1
80
[ PubMed ] Beesoon S, Bakal JA, Youngson E, Williams KP, Berzins SA, Brindle ME, Joffe AM. Décès excessifs pendant la pandémie de COVID-19 en Alberta, Canada. Numéro d’enregistrement IJID 2022 déc;5:62-67. doi: 10.1016/j.ijregi.2022.08.011. Epub 30 août 2022. PMID : 36060856 ; ID PMC : PMC9424127. https://doi.org/10.1016/j.ijregi.2022.08.011
81
Todd M, Scheeres A. Surmortalité non liée à la COVID-19 pendant la pandémie de COVID-19 à Philadelphie, Pennsylvanie, 2020-2021. Am J Public Health. Déc. 2022 ; 112(12) : 1800-1803. doi : 10.2105/AJPH.2022.307096. PMID : 36383938 ; PMCID : PMC9670212. https://doi.org/10.2105/AJPH.2022.307096
82
Karlinsky A, Kobak D. Ensemble de données sur la mortalité mondiale : Suivi de la surmortalité dans les pays pendant la pandémie de COVID-19. medRxiv [Prépublication]. 4 juin 2021 : 27 janvier 2021. doi : 10.1101/2021.01.27.21250604. Mise à jour dans : Elife. 30 juin 2021 ; 10 : PMID : 33532789 ; PMCID : PMC7852240. https://doi.org/10.1101/2021.01.27.21250604
83
Collaborateurs sur la surmortalité liée à la COVID-19. Estimation de la surmortalité due à la pandémie de COVID-19 : analyse systématique de la mortalité liée à la COVID-19, 2020-2021. Lancet. 16 avril 2022 ; 399(10334) : 1513-1536. doi : 10.1016/S0140-6736(21)02796-3. Publication en ligne du 10 mars 2022. Erratum dans : Lancet. 16 avril 2022 ; 399(10334) : 1468. PMID : 35279232 ; PMCID : PMC8912932. https://doi.org/10.1016/S0140-6736(21)02796-3
84
Msemburi W, Karlinsky A, Knutson V, Aleshin-Guendel S, Chatterji S, Wakefield J. Les estimations de l’OMS concernant la surmortalité liée à la pandémie de COVID-19. Nature. 2023 janv. ; 613(7942): 130-137. doi : 10.1038/s41586-022-05522-2. Publication en ligne du 14 déc. 2022. PMID : 36517599 ; PMCID : PMC9812776. https://doi.org/10.1038/s41586-022-05522-2
85
Shang W, Wang Y, Yuan J, Guo Z, Liu J, Liu M. Surmortalité mondiale pendant la pandémie de COVID-19 : revue systématique et méta-analyse. Vaccins (Bâle). 12 octobre 2022 ; 10(10) : 1702. doi : 10.3390/vaccines10101702. PMID : 36298567 ; PMCID : PMC9607451. https://doi.org/10.3390/vaccines10101702
86
Maiese A, Baronti A, Manetti AC, Di Paolo M, Turillazzi E, Frati P, Fineschi V. Décès après administration de vaccins contre la COVID-19 approuvés par l’EMA : un lien de causalité a-t-il été démontré ? Vaccins (Bâle). 16 févr. 2022 ; 10(2): 308. doi : 10.3390/vaccines10020308. PMID : 35214765 ; PMCID : PMC8875435. https://doi.org/10.3390/vaccines10020308
87
Sessa F, Salerno M, Esposito M, Di Nunno N, Zamboni P, Pomara C. Constatations d’autopsie et lien de causalité entre décès et vaccination contre la COVID-19 : revue systématique. J Clin Med. 15 déc. 2021 ; 10(24) : 5876. doi : 10.3390/jcm10245876. PMID : 34945172 ; PMCID : PMC8709364. https://doi.org/10.3390/jcm10245876
88
Pronker ES, Weenen TC, Commandeur H, Claassen EH, Osterhaus AD. Quantification des risques liés à la recherche et au développement de vaccins. PLoS One. 2013 ; 8(3) : e57755. doi : 10.1371/journal.pone.0057755. Publication en ligne du 20 mars 2013. PMID : 23526951 ; PMCID : PMC3603987. https://doi.org/10.1371/journal.pone.0057755
89
Falsey AR, Sobieszczyk ME, Hirsch I, Sproule S, Robb ML, Corey L, Neuzil KM, Hahn W, Hunt J, Mulligan MJ, McEvoy C, DeJesus E, Hassman M, Little SJ, Pahud BA, Durbin A, Pickrell P, Daar ES, Bush L, Solis J, Carr QO, Oyedele T, Buchbinder S, Co. et al. wden J, Vargas SL, Guerreros Benavides A, Call R, Keefer MC, Kirkpatrick BD, Pullman J, Tong T, Brewinski Isaacs M, Benkeser D, Janes HE, Nason MC, Green JA, Kelly EJ, Maaske J, Mueller N, Shoemaker K, Takas T, Marshall RP, Pangalos MN, Villafana T, Gonzalez-Lopez A; Groupe d’étude clinique AstraZeneca AZD1222. Phase 3 de sécurité et d’efficacité du vaccin Covid-19 AZD1222 (ChAdOx1 nCoV-19). N Engl J Med. 16 décembre 2021 ; 385(25) : 2348-2360. doi: 10.1056/NOMoa2105290. Epub 29 septembre 2021. PMID : 34587382 ; ID PMC : PMC8522798. https://doi.org/10.1056/NOMoa2105290
90
Matar RH, Than CA, Nakanishi H, Daniel RS, Smayra K, Sim BL, Beran A, Danoun OA. Résultats des patients présentant des événements thromboemboliques après la vaccination AstraZeneca contre le coronavirus 2019 : revue systématique et méta-analyse. Fibrinolyse des coagulations sanguines. 1er mars 2022 ; 33(2) : 90-112. doi : 10.1097/MBC.0000000000001113. PMID : 34980833 ; PMCID : PMC8815637. https://doi.org/10.1097/MBC.0000000000001113
91
Jain V, Lorgelly P. Impact de l’interruption du vaccin Oxford-AstraZeneca contre la COVID-19 sur la vaccination en Europe : analyse des différences dans les différences. Eur J Public Health. 1er août 2022 ; 32(4) : 648-654. doi : 10.1093/eurpub/ckac039. PMID : 35394507 ; PMCID : PMC9341841. https://doi.org/10.1093/eurpub/ckac039
92
Tan LJ, Koh CP, Lai SK, Poh WC, Othman MS, Hussin H. Revue systémique et recommandation pour une approche autopsique des décès après vaccination contre la COVID-19. Forensic Sci Int. 2022 Nov;340:111469. doi: 10.1016/j.forsciint.2022.111469. Publication en ligne du 20 sept. 2022. PMID: 36162300; PMCID: PMC9487151. https://doi.org/10.1016/j.forsciint.2022.111469
93
Hulscher N, Hodkinson R, Makis W, McCullough PA. Constatations d’autopsie dans les cas de myocardite mortelle induite par le vaccin contre la COVID-19. ESC Heart Fail. 14 janvier 2024. doi : 10.1002/ehf2.14680. Publication électronique avant impression. PMID : 38221509. https://doi.org/10.1002/ehf2.14680
94
Polack FP, Thomas SJ, Kitchin N, et al. Innocuité et efficacité du vaccin à ARNm BNT162b2 contre la Covid-19. N Engl J Med. 2020 ; 383(27) : 2603-2615. doi : 10.1056/NEJMoa2034577. https://doi.org/10.1056/NEJMoa2034577
95
Baden LR, El Sahly HM, Essink B, et al. Efficacité et innocuité du vaccin à ARNm-1273 contre le SARS-CoV-2. N Engl J Med. 2021 ; 384(5) : 403-416. doi : 10.1056/NEJMoa2035389. https://doi.org/10.1056/NEJMoa2035389
96
Beladiya J, Kumar A, Vasava Y, et al. Innocuité et efficacité des vaccins contre la COVID-19 : revue systématique et méta-analyse d’essais cliniques contrôlés et randomisés. Rev Med Virol. 2024 ; 34(1) : e2507. doi : 10.1002/rmv.2507. https://doi.org/10.1002/rmv.2507
97
97. Andrews CD, Parker EPK, Horne E, Walker V, Palmer T, Schaffer AL, et al. OpenSAFELY : Efficacité de la vaccination contre la COVID-19 chez les enfants et les adolescents. medRxiv. 2024 ; Disponible sur : https://www.medrxiv.org/content/early/2024/05/20/2024.05.20.24306810
98
Alessandria M, Malatesta GM, Berrino F, Donzelli A. A Critical Analysis of All-Cause Deaths during COVID-19 Vaccination in an Italian Province. Microorganisms. 2024; 12(7):1343. doi: 10.3390/microorganisms12071343. https://doi.org/10.3390/microorganisms12071343
99
Mead M, Seneff S, Wolfinger R, et al. « Vaccins » à ARNm modifié contre la COVID-19, partie 1 : Leçons tirées des essais cliniques, de la vaccination de masse et du complexe biopharmaceutique. Vaccine Theory Prac & Res. 2024 ; 3(1) : 1112-1178. doi : 10.56098/fdrasy50. https://doi.org/10.56098/fdrasy50
100
Suzumura Y. Analyse de l’association entre la vaccination contre la COVID-19 par l’ARNm BNT162b2 et les décès dans les 10 jours suivant la vaccination, en utilisant le sex-ratio au Japon. Cureus. 2023 ; 15(12) : e50144. Publié le 7 décembre 2023. doi : 10.7759/cureus.50144. https://doi.org/10.7759/cureus.50144
101
Acevedo-Whitehouse K, Bruno R. Risques potentiels pour la santé liés à la vaccination à base d’ARNm : une hypothèse. Med Hypotheses. 2023 ; 171 : 111015. doi : 10.1016/j.mehy.2023.111015. https://doi.org/10.1016/j.mehy.2023.111015
102
Igyártó BZ, Qin Z. Les vaccins ARNm-LNP : le bon, la brute et le truand ?. Front Immunol. 2024 ; 15 : 1336906. Publié le 8 février 2024. doi : 10.3389/fimmu.2024.1336906. https://doi.org/10.3389/fimmu.2024.1336906
Le Monde Diplomatique remet en question l’efficacité des confinements. Mieux vaut tard que jamais. Quant aux injections et leurs conséquences délétères, pourrons-nous lire bientôt un nouvel article à ce sujet dans leurs pages ? Pour mémoire : Covid, vaccins et science aux origines d’une défiance
« Une société hiérarchisée n’était possible que sur la base de la pauvreté et de l’ignorance. (…) Comme aucune victoire décisive n’est possible, il importe peu que la guerre se déroule bien ou mal. Il suffit que l’état de guerre existe. » George Orwell, 1984
Savez-vous ou vous rappelez-vous que pendant ou depuis l’état d’urgence…
la natalité a nettement chuté depuis 2021, et les démographes prévoient une population européenne réduite de moitié d’ici 2070 ? https://www.insee.fr/frstatistiques/7670212
près d’un million de scientifiques réputés, dépourvus de conflits d’intérêts, ont signé dès 2020 la déclaration de Great barrington contre les mesures absurdes et autoritaires des états et de l’OMS : https://gbdeclaration.org/la-declaration-de-great-barrington/
le député des Français de l’étranger Meyer Habib a écrit le 2 avril 2020 à Olivier Véran, avec copie à Emmanuel Macron et Edouard Philippe pour demander d’abroger le décret 2020-360 sur le rivotril liquide entraînant des euthanasies non consenties dans les EHPAD en l’absence des familles. Elles s’élèveront à plus de 10 000. Le Dr Hardi dénoncera la mesure sur radio Shalom le 1/04. Une audition de l’A.N. confirma les faits le 7/7/2020 https://lemediaen442.fr/pierre-chaillot-le-scandale-du-rivotril-revele-par-les-statistiques/ et https://www.youtube.com/watch?v=a3hebKg5GdM
les pays où les confinements ont été les plus sévères sont aussi ceux où la mortalité a été la plus forte (refus de soin, isolement, promiscuité, manque d’oxygène, de soleil, stress…) outre la catastrophe économique et psychologiques pour les populations. https://www.monde-diplomatique.fr/2025/03/BOULAKIA/68102
“Des régimes, des injections et des injonctions seront combinés, dés le plus jeune âge, pour produire la sorte de personnes et la sorte de croyances que les autorités considèrent désirables, et toute critique sérieuse des pouvoirs qui existent deviendra psychologiquement impossible” Bertrand Russell, prix Nobel, The Impact of Science on Society, 1944
les chiffres des effets secondaires (hors personnalités connues) doivent être multipliés au moins par 10 en raison de leur non-déclaration ou du refus des médecins de les prendre en compte. https://www.monde-diplomatique.fr/2023/04/DENOYEL/65657
aucune vaccination de masse n’a éradiqué une maladie: la variole l’a été en Afrique par l’hygiène, l’alimentation, le renforcement des défenses immunitaires, l’immunité collective et l’isolement des personnes touchées. https://boutique.arte.tv/detail/des-vaccins-et-des-hommes
« Cette sorte de pouvoirs que donne l’état d’urgence, nés de la crise, obéit à une parfaite histoire d’oppression. » Edward Snowden, 10 avril 2020 entretien avec Shane Smith, Shelter in Place
Michèle Rivasi, normalienne, agrégée de biologie, députée européenne écologiste et la première à avoir dénoncé le mensonge sur Tchernobyl, est décédée fin 2023 en allant au parlement européen à Bruxelles, avait attaqué en justice Ursula Von der Leyen pour l’opacité de la signature des accords pour plusieurs milliards d’euros entre l’UE et Pfizer, firme pour laquelle travaillait son mari. Ses derniers combats portaient sur les vaccinations, la 5G, le glyphosate. https://www.lemonde.fr/disparitions/article/2023/11/29/l-eurodeputee-ecologiste-michele-rivasi-estmorte_6202958_3382.html
Ursula von der Leyen, présidente jamais élue d’une commission européenne elle-même jamais élue, après avoir négocié les contrats Pfizer a aussi signé le mercosur permettant d’empoisonner encore davantage les populations et ruiner les agriculteurs. https://fr.businessam.be/vonder-leyen-pfizer-bourla-vaccins-ocvid-sms/
Astrazeneca et Sanofi sont derrière le beyfortus, vaccin contre la bronchiolite du nourrisson à partir de 3 jours de vie, depuis 2023, responsable de nombreux accidents (https://neosante.eu/beyfortus-alerte-rouge/)
le serment d’Hippocrate déclare: Je passerai ma vie et j’exercerai mon art dans l’innocence et la pureté (…). (…) pour l’utilité des malades, me préservant de tout méfait volontaire et corrupteur.»; le code de Nuremberg interdit toute pression pour l’administration de soins. La convention d’Oviedo interdit l’usage d’OGM pouvant affecter la descendance humaine; la charte de Munich oblige moralement la presse à ne pas conditionner sa parole à un intérêt autre que celui des populations.
Robert Malone, le pionnier des vaccins ARN, a formellement mis en garde les parents contre la vaccination ARNm des enfants : https://www.dailymotion.com/video/x86g2dl
regarder le CV professionnel et judiciaire des personnes, firmes ou administrations avant de leur faire confiance.
suivre les financements et les bénéfices des lois et décisions en répondant aux questions: quoi ? qui ? où ? quand ? combien? comment ? Pourquoi ? Sur le court, le moyen et le long terme.
écouter les victimes et témoins directs des “soins” et “médicaments” proposés, les repentis du système, et non les théoriciens et “fact checkers” financés, sans expertise ni expérience.
se demander quelles sont les conséquences des mesures présentées comme sécuritaires et salutaires en termes de libertés, démocratie, information, respect de la Nature, cohésion sociale, santé publique, qualité de vie.
couleur politique indifférente. Beaucoup ont changé de bord. Seuls comptent mots et actes.
Effets indésirables des vaccins covid :
Chez les jeunes et les adultes : troubles cardiaques graves (arrêt cardiaque, myocardite, péricardite), hémorragies cérébrales, aménorrhées, troubles menstruels, infertilité, accidents vasculaires cérébraux et morts subites, maladies auto-immunes, cancers et récidives foudroyantes de cancers avec métastases (dont de nombreux cancers de la vessie et des méninges), rechutes, nouveaux cas de SIDA, embolies pulmonaires, thromboses, thrombocytopénies, syndrome de Guillain-Barré (paralysie), accidents ischémiques transitoires, paralysies, cécité, troubles de la vision et de l’audition, fausses-couches… Chez les nouveaux-nés : troubles de la croissance, déficiences du système immunitaire, accidents vasculaires cérébraux
Une lettre ouverte de la Ligue Nationale pour la Liberté des Vaccinations soulève des préoccupations concernant les effets secondaires des vaccins à ARNm « conventionnels », une délivrance de l’Autorisation de Mise sur le Marché (AMM) d’une nouvelle génération de vaccins ARNm auto-amplifiants / réplicon ainsi que sur le classement de certains vaccins en substance vénéneuse, notamment ceux recommandés pour les femmes enceintes et ceux obligatoires pour les enfants.
La Ligue Nationale pour la Liberté des Vaccinations (LNPLV), par l’intermédiaire de son conseil Me Jean-Pierre Joseph, avocat au Barreau de Grenoble, a récemment adressé un courrier à la directrice de l’Agence Nationale de Sécurité du Médicament (ANSM) concernant les vaccins à ARN messager. Cette missive, datée du 4 mars 2025, soulève plusieurs inquiétudes sur les effets secondaires des vaccins anti-COVID-19 et l’autorisation récente d’une nouvelle génération de vaccins ARNm auto-amplifiants.
Des effets secondaires « alarmants » des vaccins ARNm conventionnels
Selon la LNPLV, le dernier rapport de pharmacovigilance publié par l’ANSM le 1ᵉʳ juin 2023 fait état de 193 934 cas d’effets indésirables déclarés pour les vaccins à ARNm Comirnaty (Pfizer) et Spikevax (Moderna). L’association s’inquiète qu’aucun rapport complémentaire n’ait été fourni depuis cette date. La lettre cite le Centre Régional de Pharmacovigilance (CRPV) de Tours, estimant que ces chiffres seraient largement sous-évalués et ne représenteraient que 1 à 10% des effets indésirables réels. En tenant compte de cette sous-déclaration, la LNPLV avance que le nombre réel d’effets secondaires pourrait atteindre environ 1,9 million de cas, dont 475 000 cas graves.
Face à ces chiffres, Me Joseph interroge l’ANSM sur les raisons qui l’ont empêchée d’exercer son pouvoir d’interdiction prévu par l’article L.5312-1 du Code de la santé publique, qui dispose que « l’agence peut interdire ces activités en cas de danger grave ou de suspicion de danger grave pour la santé humaine. »
Me Jean-Pierre Joseph, avocat (DR)
Préoccupations concernant la nouvelle génération de vaccins ARNm auto-amplifiants
La lettre aborde également l’autorisation de mise sur le marché (AMM) accordée le 14 février 2025 par la Commission européenne au vaccin KOSTAIVE, développé par ARCTURUS THERAPEUTICS et CSL. Il s’agit du premier vaccin à ARNm auto-amplifiant contre la COVID-19. La LNPLV s’inquiète de la rapidité avec laquelle cette autorisation a été délivrée pour une technologie qualifiée de nouvelle génération d’ARNm, dont la particularité est de se répliquer de manière autonome dans l’organisme après injection. L’association souligne qu’avant la mise en place du système européen de délivrance des AMM, la durée moyenne des essais cliniques pour les vaccins était d’environ 15 à 20 ans, au minimum pour des technologies classiques, alors que cette nouvelle procédure permet l’homologation en quelques mois. Me Joseph pose plusieurs questions techniques concernant la quantité d’ARN messagers qui circulera dans l’organisme après l’injection (capacité de réplication autonome des nouveaux ARNm auto-amplifiants), le risque de contagiosité de ces ARN auto-amplifiants, et les garanties concernant l’absence d’effets secondaires graves comme les thromboses, cancers, myocardites ou troubles neurodégénératifs.
Classification en « substances vénéneuses » et recommandations aux populations vulnérables
Le troisième volet de la lettre concerne la décision de l’ANSM du 7 septembre 2023 qui a porté inscription de 14 vaccins sur la liste des substances vénéneuses définies à l’article L.5132-6 du Code de la santé publique. La LNPLV s’interroge sur les raisons qui amènent l’ANSM à continuer de recommander ces injections aux femmes enceintes et aux enfants, notamment dans le cadre des 15 vaccins infantiles rendus obligatoires depuis le 1ᵉʳ janvier 2025. Cette lettre ouverte, largement diffusée puisqu’elle a été adressée en copie à de nombreux parlementaires français, des responsables européens et des avocats internationaux spécialisés, attend désormais une réponse officielle de l’ANSM sur ces questions de santé publique.
Après avoir travaillé dans l’enseignement (physiologie, biologie), Michel Georget (décédé en 2019) a concrétisé le projet qui lui tenait le plus à cœur : écrire un livre aussi exhaustif et objectif que possible sur un sujet très sensible qu’il maîtrise parfaitement : les vaccinations.
Screenshot
La qualité première de ce livre est de nous rendre lucides. L’auteur éclaire de façon incontestable certaines zones obscures de ce qui touche, en nous, l’essentiel : notre santé. Nous devrions tous lire cet ouvrage. À plus forte raison si nous avons des enfants. Mais aussi pour nous-mêmes, afin de pouvoir juger de l’opportunité de les accepter ou de les refuser et, pour les vaccinations obligatoires, obtenir la liberté totale de choix comme c’est le cas dans la plupart des pays européens. Table des matières : Chap. I : Quelques clefs pour comprendre la suite – Chap. II : Les vaccins et leur fabrication – Chap. III : À quels risques nous exposent les vaccinations? Chap. IV : Ces risques valent-ils d’être courus? – Chap. V : Le marché du vaccin, l’éthique et l’information – Chap. VI : Devons-nous réviser nos concepts ?
Des chercheurs italiens ont évalué l’impact de la vaccination contre le COVID dans une province italienne entre 2021 et 2023. L’espérance de vie des personnes vaccinées ayant reçu deux injections était diminuée de 37 %.
Une étude de cohorte conduite sur la population globale de la province de Pescara, en Italie (Flacco et al.)[1], a conclu en 2022 que la vaccination contre le COVID-19 n’avait pas entraîné de hausse de la mortalité chez les personnes vaccinées au cours des dix-huit mois qui ont suivi le déploiement des injections. Ces résultats ont été réévaluées en 2024 par des chercheurs (Alessandria et al.)[2] qui parviennent à des conclusions diamétralement opposées.
Biais de l’étude Pescara de 2022
La méthodogie de l’étude Pescara avait été critiquée en 2023 par le Comité scientifique de la fondation Allineare Sanità e Salute (Berrino et al.)[3], qui relevaient la présence de deux biais majeurs – le « biais de temps immortel » et l’effet vacciné sain[4] – conduisant à une surestimation de la mortalité chez les personnes non vaccinées et à une sous-évaluation des décès potentiellement povoqués par le vaccin dans la seconde cohorte.
L’estimation de la mortalité vaccinale dans l’étude Pescara contredit notamment les données publiques de santé anglaises, publiées mensuellement, qui démontrent que la mortalité n’a cessé d’augmenter chez les personnes vaccinées depuis 2021, le risque de décès étant proportionnel aux nombres de doses injectées (voir graphes ci-dessous commentés dans un précédent article) :
Les données mensuelles publiées dans le bulletin de l’ONS britannique montrent une augmentation progressive dans le temps du risque relatif de mortalité totale dans les groupes vaccinés par rapport aux groupes non vaccinés. Une telle augmentation pourrait également s’être produite à Pescara. Flacco et al. n’ayant pas pris en compte l’évolution des risques dans le temps, il est possible qu’ils ne l’aient pas remarquée. Le calcul des taux de mortalité moyens sur l’ensemble de la période de suivi peut en effet avoir masqué une telle tendance. Il serait souhaitable que les données d’entrée soient disponibles pour permettre une telle analyse.Berrino F, Donzelli A, Bellavite P, Malatesta G. COVID-19 vaccination and all-cause and non-COVID-19 mortality. A revaluation of a study carried out in an Italian Province. Epidemiol. Prev. 2023 Nov-Dec;47(6):374-8. https://doi.org/10.19191/ep23.6.a643.075.
Enfin, l’étude hollandaise de juin 2024[5] qui établit la persistance d’une surmortalité dans les pays occidentaux depuis la fin de la pandémie de COVID-19 contredit également l’optimisme de l’étude Pescara mais ne permet pas d’incriminer les vaccins en raison de sa méthodologie :
Un article récent a souligné la persistance d’excès importants de décès toutes causes confondues dans les années 2021 et 2022, à la suite du début de la pandémie, malgré les mesures de confinement et les programmes de vaccination. Cependant, sans distinguer les décès sur la base du statut de vaccination, il est impossible d’établir s’il existe un lien entre ces décès excessifs et les vaccinations massives contre le COVID-19.Alessandria M, Malatesta GM, Berrino F, Donzelli A. A critical analysis of all-cause deaths during COVID-19 vaccination in an Italian province. Microorganisms. 2024 Jun 30;12(7):1343. https://doi.org/10.3390/microorganisms12071343.
L’objectif de cette nouvelle étude était donc de réévaluer l’impact réel de la campagne de vaccination en termes de mortalité parmi la population de la province de Pescara (Italie), mais en éliminant le biais de temps immortel présent dans l’étude de 2022, qui utilise des périodes de suivi différentes entre personnes vaccinées et non vaccinées.
Réévaluation des données par la nouvelle étude italienne
Les auteurs ont repris les données de l’étude Pescara, actualisées en 2023 (Rosso et al.)[6].
La cohorte a été divisée en 4 groupes : non vaccinés, vaccinés une dose, deux doses ou trois/quatre doses. La période de vaccination couvrait le début de la campagne jusqu’au 31 décembre 2022. Le suivi de la mortalité a porté sur la période allant du 1er janvier 2021 et le 15 février 2023.
La période d’observation a duré 775 jours en fonction des dates de début et de fin spécifiques à chaque cohorte :
non vacciné : 258 jours ;
vacciné 1 dose : 61 jours ;
vacciné 2 doses : 247 jours ;
vacciné 3/4 doses : 400 jours.
Les auteurs ont suivi l’évolution de la mortalité pour chacun des quatre groupes. Pour chaque dose, ils ont comparé le risque de décès toutes causes entre personnes vaccinées vs non vaccinées, ainsi que le risque de décès spécifique aux personnes présentant une ou plusieurs comorbidités en fonction de leur statut vaccinal. Les comorbidités étudiées étaient l’hypertension, le diabète, le cancer, la BPCO, une maladie rénale, une infection COVID.
Les personnes vaccinées avaient un risque de décès global accru par rapport aux personnes non vaccinées quel que soit le nombre de doses, hormis pour les doses de rappel. Ce risque était beaucoup plus élevé que celui retrouvé dans l’étude Pescara : + 140 % vs 40 % après la dose 1, + 98 % vs + 36 % après la dose 2, – 1 % vs – 78 % après 3 doses ou plus.
Les personnes vaccinées présentant au moins une comobordité avaient un risque de décès accru par rapport aux personnes sans comorbidités avec le même schéma vaccinal, ce qui suggère que le vaccin n’est pas efficace 100 % contre les formes à graves, voire qu’il exacerbe les comorbidités chez les personnes à risque.
Impact de la vaccination sur l’espérance de vie
Les auteurs ont également évalué l’impact de la vaccination sur l’espérance de vie en fonction du nombre de doses reçues, au moyen de deux indicateurs :
la durée de survie moyenne restreinte (RMST, restricted mean survival time), qui correspond au délai écoulé entre l’injection ou le début de la période de suivi et le décès ;
la durée de survie moyenne restreinte perdue (RMTL, restricted mean time loss), qui représente les jours de vie perdus par rapport à la durée de la période de suivi.
La différence entre le RMTL des personnes vaccinées vs non vaccinées correspond au nombre de jours de vie perdus dans la population vaccinée. Le ratio entre ces deux RMTL correspond à l’espérance de vie perdue.
Résultat
Pour les personnes vaccinées à deux doses, la durée de survie était de 728,9 jours vs 731,6 jours pour les personnes non vaccinées sur la période de suivi de 739 jours, ce qui représente une différence d’espérance de vie (RMST) de 2,71 jours. La perte d’espérance de vie (RMTL) était de 739 – 728,9 = 10,1 jours pour les personnes vaccinées et de 739 – 731,6 = 7,4 jours pour les personnes non vaccinées, ce qui représente une perte égale à 1,37 fois celle des non-vaccinés.
Pour les personnes vaccinées à trois doses ou plus, le RMST était de 574,4 jours vs 573,7 jours sur la période de suivi de 579 jours, ce qui représente une perte d’espérance de vie de 0,764 jour pour les personnes vaccinées, équivalant à 1,17 fois celle des non-vaccinées.
Interprétation
Concernant le RMTL, cela signifie que les sujets vaccinés avec trois doses ou plus ont perdu 17 % d’espérance de vie par rapport à la population non vaccinée au cours de la période de suivi. Pour les personnes vaccinées à deux doses, la perte d’espérance de vie par rapport à une personne non vaccinée atteint 37 %.
Concernant le RMST, la différence entre l’espérance de vie des personnes vaccinées vs non-vaccinées n’est que de 2,71 jours à deux doses et 0,764 jour à trois doses ou plus, mais pour des périodes de suivi de 739 jours et 579 jours. Ces chiffres semblent anecdotiques, mais lorsqu’on les extrapole à l’espérance de vie de la population d’étude (82,6 ans), cela représente en réalité une différence d’espérance de vie d’environ 3,6 mois et 1,31 mois, voire de 4 mois si on se réfère à l’espérance de vie de 83,6 ans qui prévalait en 2019, avant la pandémie de COVID.
Conclusion des auteurs
Nous avons constaté que les risques de décès toutes causes confondues étaient encore plus élevés chez les personnes vaccinées avec une et deux doses par rapport aux personnes non vaccinées et que les doses de rappel étaient inefficaces. Nous avons également constaté une perte légère mais statistiquement significative de l’espérance de vie des personnes vaccinées avec 2 doses ou avec 3 ou 4 doses.
Sous-estimation des effets secondaires de la vaccination
Selon les auteurs, plusieurs biais subsistent qui pourraient conduire à une sous-estimation des effets indésirables des injections et de la létalité des doses de rappel.
L’effet de moisson (harvesting effect)
La vaccination a eu un impact significatif sur les décès toutes causes des personnes vaccinées avec 1 ou 2 doses mais pas sur celles ayant reçu 3 doses ou plus. En supposant que la vaccination a réduit les décès liés au COVID-19 – ce qui semble le cas puisque le taux de décès COVID est le plus élevé dans la population non vaccinée –, cette réduction a donc été contrebalancée par une augmentation des décès dus à d’autres causes, qui incluent les effets secondaires des injections.
Le biais de calendrier
Les rappels ont commencé en juillet 2021, lorsque la vague pandémique était terminée. A contrario, le suivi des personnes non vaccinées et des vaccinées à une dose a commencé en janvier 2021, lorsque le risque de décès COVID-19 et de décès toutes causes était significativement plus élevé, ce qui conduit à sous-estimer les décès toutes causes des personnes vaccinées avec trois doses ou plus.
Le biais de fenêtre de comptage de cas
Ce biais consiste à classer comme « non vaccinées » les personnes vaccinées durant les 14 jours suivant une injection. Il a été exploité dans tous les pays pour simuler une pandémie de non vaccinés, dont les fuites du RKI allemand ont démontré qu’elle n’avait jamais existé, et réduire artificiellement la part des décès touchant les personnes vaccinées pour susciter l’idée que le vaccin réduit la mortalité. Or une étude réalisée avec les données de l’essai clinique du vaccin Pfizer (Fung et al.)[7] a montré qu’un vaccin inefficace pouvait sembler efficace à 48 %, en raison de ce changement de 14 jours dans la prise en compte du statut vaccinal. Cette règle a été notamment utilisée par les CDC, l’Office national des statistiques anglais et l’Institut supérieur de la santé italien.
Le biais du vacciné sain
En 2021, en Italie, le public prioritaire de la vaccination a inclus des personnes réputées en bonne santé (professionnels de santé, policiers, militaires, enseignants…). Selon les auteurs, ce biais aujourd’hui largement documenté est extrêmement puissant, comme l’a démontré l’analyse[8],[9] d’une étude nationale sur la mortalité en fonction du statut vaccinal contre le COVID en République tchèque. Il a donc très probablement influencé les comportements vaccinaux dans cette cohorte, y compris celui des personnes fragiles à un stade avancé de leur maladie, pour qui la vaccination n’est souvent même pas envisagée (biais d’exclusion de la fragilité).
Conclusion
Cette étude confirme la tendance observée dans l’ensemble des pays où les gouvernements ont massivement injecté leur population avec des vaccins ARNm dont ils savaient qu’ils étaient susceptibles d’engendre un nombre inacceptable d’effets indésirables. Pour une raison qui reste à déterminer, ils ont choisi d’endosser ce risque juridique à la place des laboratoires en leur assurant une immunité totale en cas de blessures.
Cette étude en revanche la première, à notre connaissance, qui évalue l’impact de la vaccination en termes de perte d’espérance de vie. Selon les auteurs, il s’agit d’une simple extrapolation « faite dans le seul but de donner au lecteur une idée de l’ordre de grandeur du nombre de jours de vie perdus » qui ne prétend pas être une « prédiction réaliste, car elle présupposerait que les conditions de santé soient invariantes dans le temps, une hypothèse très difficile à réaliser ». Le poids de cette perte d’espérance de vie n’est en revanche pas quantifiable pour les parents qui ont perdu un enfant qui n’aurait jamais dû se faire vacciner contre le COVID-19.
Enfin, elle redonne de la visibilité à l’étude hollandaise publiée en juin dernier dans laquelle les auteurs s’étonnent que la surmortalité observée pendant la pandémie se soit aggravée depuis la campagne de vaccination et qu’elle ne se soit toujours pas résorbée à ce jour. L’étude a été rétractée en toute discrétion 11 jours après sa mise en ligne après avoir provoqué un véritable séisme dans la presse internationale. Est-on autorisé à se demander « pourquoi ? » et à s’interroger sur les raisons du silence indécent des médias français ?
Références
[1] Flacco ME, Acuti Martellucci C, Soldato G, Di Martino G, Carota R, De Benedictis M, et al. COVID-19 vaccination did not increase the risk of potentially related serious adverse events: 18-month cohort study in an Italian province. Vaccines. 2022 Dec 23;11(1):31. https://doi.org/10.3390/vaccines11010031.
[2] Alessandria M, Malatesta GM, Berrino F, Donzelli A. A critical analysis of all-cause deaths during COVID-19 vaccination in an Italian province. Microorganisms. 2024 Jun 30;12(7):1343. https://doi.org/10.3390/microorganisms12071343.
[3] Berrino F, Donzelli A, Bellavite P, Malatesta G. COVID-19 vaccination and all-cause and non-COVID-19 mortality. A revaluation of a study carried out in an Italian Province. Epidemiol. Prev. 2023 Nov-Dec;47(6):374-8. https://doi.org/10.19191/ep23.6.a643.075.
[4] Un biais de temps immortel se produit lorsque les participants à un essai clinique ne sont pas évalués sur la même période selon qu’ils appartiennent au groupe placebo qui reçoit un traitement ou au groupe placebo, en raison d’un décalage entre les dates de recrutements ou de durées de suivi différentes, en cas de décès par exemple. Ce biais est fréquent dans les études de cohorte.
Le biais dit « du patient en bonne santé » ou du « vacciné sain » décrit le fait que dans une étude observationnelle en évaluation, d’une vaccination par exemple, ces médicaments seront prioritairement administrés aux enfants en pleine santé. Les personnes fragiles ou malades seront alors reléguées dans le groupe contrôle etet la fréquence des effets secondaires sera artificiellement diminuée dans le groupe vacciné (voir le site de l’Amsib pour une explication plus détaillée).
[5] Mostert S, Hoogland M, Huibers M, Kaspers G. Excess mortality across countries in the Western World since the COVID-19 pandemic: ‘Our World in Data’ estimates of January 2020 to December 2022. BMJ Public Health 2024;2:e000282. https://doi.org/10.1136/bmjph-2023-000282.
[6] Rosso A, Flacco ME, Soldato G, Di Martino G, Acuti Martellucci C, Carota R, et al. COVID-19 vaccination effectiveness in the general population of an Italian province: two years of follow-up. Vaccines (Basel). 2023 Aug 4;11(8):1325. https://doi.org/10.3390/vaccines11081325.
[8] Furst T, Straka R, Janosek J. Healthy vaccinee effect: A bias not to be forgotten in observational studies on COVID-19 vaccine effectiveness. Pol Arch Intern Med. 2024 Feb 28;134(2):16634. https://doi.org/10.20452/pamw.16634.
[9] Fürst T, Bazalová A, Fryčák T, Janošek J. Does the healthy vaccinee bias rule them all? Association of COVID-19 vaccination status and all-cause mortality from an analysis of data from 2.2 million individual health records. Int J Infect Dis. 2024 May;142:106976. https://doi.org/10.1016/j.ijid.2024.02.019.
D’où vient l’idéologie transgenre ? Comment s’est-elle propagée aussi vite dans nos sociétés ? Qui finance cet engrenage… qui broie tous ceux qui tombent dans le piège ? Ce film raconte l’incroyable suite d’événements, planifiés et orchestrés. Une poignée de milliardaires LGBT et alliés « progressistes » aux USA ont mis en commun leurs moyens financiers colossaux. Objectif : détruire le « dimorphisme sexuel humain » afin d’infuser l’idéologie transgenre dans toutes les strates de la société. Au point d’être enseignée aujourd’hui à l’école, et d’impacter la vie quotidienne de millions de personnes dans de nombreux pays.
L’affaire portée par le professeur Frajese devant la Cour de justice de l’Union européenne a connu une issue surprenante. Non pas parce que la demande de révocation de l’autorisation de mise sur le marché des vaccins, défendue par les avocats Olga Milanese (Umanità e Ragione) et Andrea Montanari (Eunomis), a été rejetée, mais en raison des décisions confirmées par l’arrêt.
En effet, selon la Cour, une prescription médicale était nécessaire pour administrer les vaccins anti-Covid. Mais il y a plus : les médecins auraient pu choisir de les administrer ou non et même les déconseiller, à tel point que l’éventuelle responsabilité civile et pénale des médecins est imputable au cas d’espèce. Les motivations de la Cour pourraient démanteler la base des procédures disciplinaires et pénales engagées contre les médecins qui se sont opposés aux vaccinations et attribuer une responsabilité sérieuse aux blouses blanches qui ont vacciné “sans si et sans mais”, favorisant ainsi le risque de provoquer des effets indésirables. Nous en avons parlé avec l’avocate Olga Milanese.
Madame l’avocate, la Cour a déclaré le professeur Frajese inintéressé à introduire un recours devant la Cour européenne pour faire annuler les autorisations de mise sur le marché des vaccins Covid. Pouvez-vous nous expliquer brièvement les motifs de cet arrêt ?
“Je dois d’abord dire que nous savions parfaitement qu’il était peu probable que la Cour nous permette de surmonter l’obstacle de la recevabilité du recours, mais nous avons décidé d’essayer quand même, à la fois parce que nos arguments étaient très solides et parce qu’une fois que le délai de recours contre les autorisations de mise sur le marché aurait expiré, il ne serait plus possible d’emprunter la voie de l’annulation. Les recours devant le Tribunal de l’UE sont soumis à des filtres très stricts. Pour contester un acte de la Commission européenne, il faut démontrer l’existence d’un intérêt juridique qualifié, d’une position particulière qui justifie l’intérêt de demander son annulation”.
Signification ?
“En d’autres termes, il faut démontrer que l’annulation de l’acte est susceptible de produire des conséquences juridiques pour la personne qui agit, que l’issue positive de l’affaire est susceptible de procurer un avantage à la partie qui introduit le recours. C’est pourquoi, à notre avis, le recours en annulation des actes autorisant la mise sur le marché des vaccins Covid-19 ne pouvait être introduit que par un médecin. En effet, l’acte médical de vaccination est une conséquence directe des autorisations litigieuses ; l’objet même des actes d’autorisation est de permettre l’utilisation des produits autorisés sur le territoire de l’Union européenne, dans le respect des prescriptions qui y figurent et donc, en l’espèce, l’administration du médicament. À tel point que les annexes mêmes des décisions d’exécution exigent, pour l’administration du produit autorisé, une prescription médicale, qui est précisément une activité confiée aux seuls médecins vaccinateurs.
Afin de convaincre le Tribunal de ne pas s’arrêter au filtre de la recevabilité et d’analyser le bien-fondé des questions posées, nous avons souligné que les décisions attaquées de la Commission, et donc la commercialisation des vaccins Covid, impliquent l’obligation pour tous les médecins vaccinateurs de devoir considérer les risques et les bénéfices du médicament, dans l’exercice de leurs fonctions spécifiques, évaluation qu’ils ne seraient en revanche pas tenus de faire en cas d’annulation des actes d’autorisation et de retrait consécutif du produit du marché. D’où l’intérêt pour le professeur Frajese de porter l’affaire devant le Tribunal de l’UE en raison des implications directes des décisions attaquées sur les activités des médecins et leurs choix professionnels.
Nous avons également rappelé à la Cour le grand problème de l’absence d’instruments capables d’inciter les organismes de réglementation à effectuer une vérification effective et non formelle de la sécurité des produits que le médecin est appelé à évaluer et à administrer, et le problème tout aussi énorme de l’absence de recours juridictionnels, à l’exception de celui que nous avons choisi, qui peuvent être activés pour contester et/ou remettre en cause les actes autorisant la commercialisation des médicaments de la classe Covid-19.
La Cour n’a pas voulu reconnaître l’existence d’un intérêt spécifique du corps médical à demander l’annulation des actes autorisant la commercialisation des médicaments, arguant que les seules personnes habilitées à introduire un tel recours sont les destinataires des actes eux-mêmes, c’est-à-dire les sociétés pharmaceutiques, qui n’introduiraient évidemment jamais un tel recours.
Il va sans dire que cela revient à affirmer l’inopposabilité substantielle des décisions de la Commission européenne dans un secteur très important, celui de la santé, qui est par ailleurs exposé à d’énormes conflits d’intérêts, puisqu’il s’agit de décisions permettant la mise sur le marché de produits destinés à la prévention ou au traitement des personnes. Tout cela en l’absence non seulement d’un contrôle tiers et impartial sur la sécurité du produit, mais d’un contrôle quelconque, ce qui peut être démontré”.
Malgré l’issue défavorable de l’arrêt, la Cour a déclaré que les vaccins anti-Covid devaient être administrés sur ordonnance, sous réserve de la liberté du médecin de les déconseiller. N’y voyez-vous pas là un échec retentissant de la part de la Cour ?
“En fait, nous avions espéré que notre reconstitution en temps utile des motifs de l’intérêt à agir de la requérante nécessiterait une analyse du bien-fondé des questions que nous avions soulevées, et c’est ce qui s’est passé.
Il n’y avait que deux possibilités : soit confirmer l’incapacité du médecin à évaluer les vaccins Covid, en choisissant de les administrer ou non et à quel moment (ce qui aurait dû conduire à la reconnaissance de son intérêt spécifique et personnel à demander l’annulation des mesures de commercialisation), soit déclarer sa liberté d’évaluation et de choix afin de nier son intérêt à porter l’affaire devant la Cour européenne.
Dans les deux cas, nous aurions obtenu un arrêt important, et c’est ce que nous avons fait. Certes, nous espérions un examen du bien-fondé de notre demande d’annulation des autorisations (avec pour conséquence le retrait du marché des produits en cause), compte tenu également de l’immense travail accompli pour démontrer l’absence des conditions préalables à l’octroi des autorisations en collectant, traduisant, numérotant et collationnant toutes les études scientifiques attestant de l’absence de sécurité de ces produits, mais le résultat “secondaire” obtenu n’est en rien insignifiant”.
Pourquoi, de votre point de vue, était-il si important d’attirer l’attention sur la commercialisation des produits anti-Covid ?
“Je crois que les documents officiels montrent clairement que le processus d’autorisation a été mené en violation non seulement de la réglementation européenne, mais aussi des règles les plus banales de prudence, de précaution et de bon sens.
J’en ai également longuement discuté avec le professeur Marco Cosentino (médecin, professeur de pharmacologie à la faculté de médecine de l’université d’Insubrie, où il dirige le Centre de recherche en pharmacologie médicale, ndlr) dans certaines de nos interventions précédentes. Cependant, le sujet est tellement technique qu’il est difficile à comprendre par les professionnels eux-mêmes, alors imaginez la difficulté de le rendre intelligible dans les salles d’audience ou pour les gens ordinaires. Et pourtant, elle est fondamentale, car si nous pouvions comprendre que la législation communautaire, avant même la législation nationale, théoriquement placée à la présidence du processus de vérification et d’autorisation de mise sur le marché des médicaments n’est pas respectée et n’offre donc aucune garantie de contrôle, alors la justice commencerait elle aussi à avancer avec plus de courage dans la seule direction possible”.
Quels seront les effets possibles de l’arrêt ?
La Cour a dû confirmer, même brièvement, que les décisions d’autorisation de mise sur le marché prises par la Commission “n’entraînent aucune obligation pour les médecins de prescrire et d’administrer ces vaccins à leurs patients”. Elle a réaffirmé le principe fondamental du droit à la liberté de traitement et au choix du traitement le plus approprié, le plus sûr et le plus efficace par le médecin, en conscience, dans le cas concret et dans l’intérêt exclusif de la santé de son patient. Ce passage est extraordinairement important parce qu’il démonte définitivement les accusations qui ont été portées en Italie, tant devant les tribunaux que dans le cadre de procédures disciplinaires, contre tous les médecins qui ont déconseillé à leurs patients la vaccination Covid ou qui ont refusé de la promouvoir, en restituant au médecin sa pleine liberté de soins.
En outre, elle confirme qu’il existe une responsabilité spécifique des médecins vaccinateurs et des ASL qui ont administré le médicament sans en évaluer correctement l’opportunité, les risques et la sécurité dans le cas spécifique du patient traité. Plus généralement, la Cour précise que “si la délivrance d’une AMM pour un vaccin est une condition préalable au droit de son titulaire de mettre ce vaccin sur le marché dans chaque État membre, cette AMM n’entraîne en principe aucune obligation à la charge des patients ou des médecins vaccinateurs”, mais surtout elle confirme qu’”il ressort des annexes des décisions attaquées qu’une prescription médicale est nécessaire pour l’administration des vaccins en cause”.
C’est ce que nous avons toujours affirmé dans les recours en faveur des travailleurs suspendus, qui avaient refusé de se faire vacciner également en raison de l’absence d’une prescription médicale spécifique, alors que dans de nombreux cas ils en avaient eux-mêmes fait la demande auprès de leur médecin traitant. La prescription n’a jamais été délivrée pour aucune des millions de doses administrées aux Italiens, rendant toutes ces administrations contra legem (ndlr contre la loi – exemption valable pour ceux qui ne voulaient pas se faire vacciner), avec ce qui s’ensuit sur le plan juridique de l’illégitimité des dispositions réglementaires imposant l’obligation et de l’illégitimité de “l’acte médical” de l’administration spécifique”.
Parlons du “bouclier pénal” pour les médecins. Quelles responsabilités pourraient être attribuées aux médecins vaccinateurs ?
“Les arrêts de la Cour peuvent influencer les procédures civiles et pénales pour la réparation des dommages (biologiques, moraux et financiers) causés aux personnes soumises à de tels traitements médicamenteux, ayant été administrés – pour la responsabilité pour faute professionnelle supportée par les ASL et les médecins vaccinateurs (du vaccin HUB ou MMG) – “en violation de la loi” pour l’absence de prescription médicale préalable (prescription limitative répétable, cd RRL). Pour expliquer en termes compréhensibles pour ceux qui ne sont pas dans l’industrie, le bouclier pénal ne fonctionne que si le traitement médical est administré conformément aux indications fournies par les actes d’autorisation qui, dans ce cas, ont été ignorés et pas seulement pour le manque d’évaluation médicale attentive et adéquate de chaque patient hésitant dans l’acte formel de la prescription. Le moment et le nombre de doses administrées n’étaient très souvent pas conformes aux indications en vigueur au moment des différentes administrations et cela empêche le fonctionnement du bouclier pénal”.
Après l’arrêt de la Cour, l’ensemble de la campagne de vaccination anti-Covid peut-il être remis en cause, y compris par le biais de la commission d’enquête ?
“Je pense que la commission d’enquête dispose déjà de nombreux éléments pour remettre en question l’ensemble de la campagne de vaccination et j’espère sincèrement qu’une table de discussion sérieuse sera ouverte à ce sujet. Il ne fait aucun doute que le contenu de l’arrêt est utile, tout comme l’analyse effectuée dans notre recours avec les nombreux documents qui l’accompagnent, documents que nous ne manquerons pas de transmettre à la Commission”.
Les arrêts de la Cour de justice de l’Union européenne s’imposent également aux juges nationaux auxquels la même question est soumise : quelles sont les perspectives pour les affaires encore pendantes, y compris celles impliquant des médecins suspendus et/ou radiés pendant la période Covid ?
“Comme indiqué plus haut, les principes affirmés dans cet arrêt ne peuvent être ignorés par les juridictions nationales, mais il est important qu’ils soient rappelés de manière correcte et pertinente. Cela dépendra de la manière dont les appels dans les arrêts seront formulés, et des motifs et arguments avancés pour soutenir l’illégalité des mesures prises. Sans aucun doute, il sera crucial d’avoir soulevé la question de la violation du droit communautaire et d’avoir ainsi mis en évidence le conflit entre la législation nationale et la législation européenne. La CJUE a rappelé dans plusieurs passages de l’arrêt qu’il appartenait aux médecins d’apprécier dans le cas concret l’opportunité d’administrer les vaccins Covid-19, en confirmant la nécessité d’une prescription à cet effet, de sorte que la règle nationale contraire à ces principes et, avant même, aux protocoles d’administration contenus dans les actes d’autorisation, rencontre la limite de l’inapplicabilité en tant qu’illégale”.
La Fnomceo a affirmé à plusieurs reprises, même avant l’ère Covid, qu’un médecin a le devoir de promouvoir activement les campagnes de vaccination et qu’il ne peut pas, sous peine de violer le code de déontologie, “déconseiller” les vaccins, quels qu’ils soient. Pourrait-il changer de position aujourd’hui ?
“Oui, il devrait ! Et il devrait revenir sur ses décisions antérieures en assumant la responsabilité de la “politique” qu’il a voulu adopter et en réparant les dommages causés aux nombreux médecins injustement harcelés par des dispositions absurdes qui sont clairement en contradiction avec les indications thérapeutiques des actes d’autorisation communautaire, avec le principe de précaution et avec le serment d’Hippocrate”.
Christine Cotton, bio-statisticienne a déposé une plainte pénale pour tromperie et administration d’une substance sans consentement contre la has, l’ansm , l’Académie de médecine, Olivier Véran, pfizer, BioNTech_Group.
Voir également : Florence Bessy, avocate spécialisée en dommages corporels, défend les soignants suspendus de la crise Covid en lançant des recours contre leurs employeurs et l’État pour obtenir des indemnités. Elle a aussi déposé un recours contre l’ANSM pour retirer les vaccins anti-Covid du marché français, aux côtés de la biostatisticienne Christine Cotton et d’autres collectifs de victimes. Lors des Rencontres citoyennes internationales à Vierzon, elle rappelle que les soignants suspendus ont jusqu’au 31 décembre 2025 pour entamer un recours contre l’État.
Nous avions déjà pointé du doigt les nombreuses pathologies liées aux vaccins COVID[1]. Désormais, 263 études revues par les pairs confirment que la protéine spike, composant clé des vaccins à ARNm, agit comme un pathogène indépendant, capable de provoquer des dysfonctionnements cellulaires et systémiques graves. Cette situation est d’autant plus préoccupante que la production de spike lors des injections est plus intense chez les jeunes, et que la quantité produite varie considérablement d’un individu à l’autre, rendant le vaccin encore plus toxique… ou encore moins efficace.
Protéine spike de la COVID‑19 : pathogénicité
Bibliothèque de recherche compilée par le Dr Martin Wucher, MSc Dent Sc (équivalent DDS), Erik Sass, et al.
DOI : 10.5281/zenodo.14269255
Dernière mise à jour : 3 décembre 2024.
À l’origine partie de l’enveloppe externe du virus SARS‑CoV‑2, où elle fonctionne comme une « clé » permettant de « déverrouiller » (infecter) les cellules, la protéine spike est également produite en grande quantité par les vaccins à ARNm, déclenchant une réponse immunitaire de courte durée sous forme d’anticorps. Cependant, un nombre croissant de preuves montre que la protéine spike est nocive en elle‑même, indépendamment du reste du virus.
La liste regroupe 263 études scientifiques évaluées par des pairs confirmant que la protéine spike est hautement pathogène par elle‑même. La plupart des études in vitro citées ici ont utilisé des protéines spike recombinantes ou des protéines spike dans des vecteurs pseudoviraux, produisant des effets pathologiques non dépendants de la machinerie virale du SARS‑CoV‑2.
Les études sont regroupées par pathologie en 32 sections, incluant les tissus et systèmes organiques affectés, les mécanismes, et les preuves issues de la pathologie clinique. Étant donné que ces domaines se chevauchent, certaines études parmi les 263 peuvent apparaître dans plusieurs sections.
Il convient de souligner qu’un tel florilège d’études distingue ce vaccin de la majorité des autres vaccins et devrait, d’un point de vue scientifique et rationnel, conduire à l’abandon de ce procédé dans le cadre d’une utilisation indiscriminée. Ce mécanisme, basé sur la production de l’immunogène par un procédé génétique directement chez le vacciné, représente une approche totalement inédite dans l’histoire des vaccins. S’écartant radicalement des méthodes traditionnelles, il s’avère que l’expérience acquise dans le domaine de l’immunothérapie du cancer était largement insuffisante pour en comprendre ou anticiper les implications.
Catégories
Général (20)
ACE2 (18)
Propriétés amyloïdes, de type prion (12)
Auto‑immune (2)
Pression artérielle/hypertension (2)
CD147 (13)
Perméabilité de la membrane cellulaire, dysfonctionnement de la barrière (13)
Acevedo‑Whitehouse K and R Bruno, “Potential health risks of mRNA‑based vaccine therapy: A hypothesis,” Med. Hypotheses 2023, 171: 111015. doi: https://doi.org/10.1016/j.mehy.2023.111015
Almehdi AM et al., “SARS‑CoV‑2 Spike Protein: Pathogenesis, Vaccines, and Potential Therapies,” Infection 2021, 49, 5: 855–876. doi: https://doi.org/10.1007/s15010‑021‑01677‑8
Baldari CT et al., “Emerging Roles of SARS‑CoV‑2 Spike‑ACE2 in Immune Evasion and Pathogenesis,” Trends Immunol. 2023, 44, 6. doi: https://doi.org/10.1016/j.it.2023.04.001
Cosentino M and Franca Marino, “Understanding the Pharmacology of COVID- 19 mRNA Vaccines: Playing Dice with the Spike?” Int. J. Mol. Sci. 2022, 23, 18: 10881. doi: https://doi.org/10.3390/ijms231810881
Gussow AB et al., “Genomic Determinants of Pathogenicity in SARS‑CoV‑2 and Other Human Coronaviruses,” PNAS 117, 2020, 26: 15193–15199. doi: https://doi.org/10.1073/pnas.2008176117
Lehmann KJ, “Impact of SARS‑CoV‑2 Spikes on Safety of Spike‑Based COVID‑19 Vaccinations,” Immunome Res. 2024, 20, 2: 1000267. doi: 10.35248/1745‑7580.24.20.267
Lehmann KJ, “Suspected Causes of the Specific Intolerance Profile of Spike‑Based Covid‑19 Vaccines,” Med. Res. Arch 2024, 12, 9. doi: 10.18103/mra.v12i9.5704
Lesgard JF et al., “Toxicity of SARS‑CoV‑2 Spike Protein from the Virus and Produced from COVID‑19 mRNA or Adenoviral DNA Vaccines,” Arch Microbiol Immun 2023, 7, 3: 121- 138. doi: 10.26502/ami.936500110
Letarov AV et al., “Free SARS‑CoV‑2 Spike Protein S1 Particles May Play a Role in the Pathogenesis of COVID‑19 Infection,” Biochemistry (Moscow) 2021, 86, 257–261. doi: https://doi.org/10.1134/S0006297921030032
Nuovo JG et al., “Endothelial Cell Damage Is the Central Part of COVID‑19 and a Mouse Model Induced by Injection of the S1 Subunit of the Spike Protein,” Ann. Diagn. Pathol. 2021, 51, 151682. doi: https://doi.org/10.1016/j.anndiagpath.2020.151682
Pallas RM, “Innate and adaptative immune mechanisms of COVID‑19 vaccines. Serious adverse events associated with SARS‑CoV‑2 vaccination: A systematic review,” Vacunas (English ed.) 2024, 25, 2: 285.e1‑285.e94. doi: https://doi.org/10.1016/j.vacune.2024.05.002
Parry PL et al., “‘Spikeopathy’: COVID‑19 Spike Protein Is Pathogenic, from Both Virus and Vaccine mRNA,” Biomedicine 2023, 11, 8: 2287. doi: https://doi.org/10.3390/biomedicines11082287
Saadi F et al., “Spike glycoprotein is central to coronavirus pathogenesis‑parallel between m‑CoV and SARS‑CoV‑2,” Ann Neurosci. 2021, 28 (3‑4): 201–218. doi: https://doi.org/10.1177/09727531211023755
Swank Z, et al. “Persistent Circulating Severe Acute Respiratory Syndrome Coronavirus 2 Spike Is Associated With Post‑acute Coronavirus Disease 2019 Sequelae,” Clin. Infect. Dis 2023, 76, 3: e487– e490. doi: https://doi.org/10.1093/cid/ciac722
Theoharides TC, “Could SARS‑CoV‑2 Spike Protein Be Responsible for Long‑COVID Syndrome?” Mol. Neurobiol. 2022, 59, 3: 1850–1861, doi: https://doi.org/10.1007/s12035‑021‑02696‑0
Theoharides TC and P. Conti, “Be Aware of SARS‑CoV‑2 Spike Protein: There Is More Than Meets the Eye,” J Biol Reg Homeostat Agents 2021, 35, 3: 833–838 doi: 10.23812/THEO_EDIT_3_21
Tyrkalska SD et al., “Differential proinflammatory activities of spike proteins of SARS‑CoV‑2 variants of concern,” Sci. Adv. 2022, 8, 37: eabo0732. doi: https://doi.org/10.1126/sciadv.abo0732
B. ACE2
Aboudounya MM and RJ Heads, “COVID‑19 and Toll‑Like Receptor 4 (TLR4): SARS‑CoV‑2 May Bind and Activate TLR4 to Increase ACE2 Expression, Facilitating Entry and Causing Hyperinflammation,” Mediators Inffamm. 2021, 8874339. doi: https://doi.org/10.1155/2021/8874339
Aksenova AY et al., “The increased amyloidogenicity of Spike RBD and pH‑dependent binding to ACE2 may contribute to the transmissibility and pathogenic properties of SARS‑CoV‑2 omicron as suggested by in silico study,” Int J Mol Sci. 2022, 23, 21: 13502. doi: https://doi.org/10.3390/ijms232113502
Angeli F et al., “COVID‑19, vaccines and deficiency of ACE2 and other angiotensinases. Closing the loop on the ‘Spike effect’,” Eur J. Intern. Med. 2022, 103: 23–28. doi: https://doi.org/10.1016/j.ejim.2022.06.015
Baldari CT et al., “Emerging Roles of SARS‑CoV‑2 Spike‑ACE2 in Immune Evasion and Pathogenesis,” Trends Immunol. 2023, 44, 6. doi: https://doi.org/10.1016/j.it.2023.04.001
Devaux CA and L. Camoin‑Jau, “Molecular mimicry of the viral spike in the SARS‑CoV‑2 vaccine possibly triggers transient dysregulation of ACE2, leading to vascular and coagulation dysfunction similar to SARS‑CoV‑2 infection,” Viruses 2023, 15, 5: 1045. doi: https://doi.org/10.3390/v15051045
Gao X et al., “Spike‑Mediated ACE2 Down‑Regulation Was Involved in the Pathogenesis of SARS‑CoV- 2 Infection,” J. Infect. 2022, 85, 4: 418–427. doi: https://doi.org/10.1016/j.jinf.2022.06.030
Kato Y et al., “TRPC3‑Nox2 Protein Complex Formation Increases the Risk of SARS‑CoV‑2 Spike Protein‑Induced Cardiomyocyte Dysfunction through ACE2 Upregulation,” Int. J. Mol. Sci. 2023, 24, 1:102. doi: https://doi.org/10.3390/ijms24010102
Ken W et al., “Low dose radiation therapy attenuates ACE2 depression and inflammatory cytokines induction by COVID‑19 viral spike protein in human bronchial epithelial cells,” Int J Radiat Biol. 2022, 98, 10: 1532‑1541. doi: https://doi.org/10.1080/09553002.2022.2055806
Lei Y et al., “SARS‑CoV‑2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2,” Circulation Research 2021, 128, 9: 1323–1326. doi: https://doi.org/10.1161/CIRCRESAHA.121.318902
Lu J and PD Sun, “High affinity binding of SARS‑CoV‑2 spike protein enhances ACE2 carboxypeptidase activity,” J. Biol. Chem 2020, 295, 52: p18579‑18588. doi: 10.1074/jbc.RA120.015303
Maeda Y et al., “Differential Ability of Spike Protein of SARS‑CoV‑2 Variants to Downregulate ACE2,” Int. J. Mol. Sci. 2024, 25, 2: 1353. doi: https://doi.org/10.3390/ijms25021353
Magro N et al., “Disruption of the blood‑brain barrier is correlated with spike endocytosis by ACE2 + endothelia in the CNS microvasculature in fatal COVID‑19. Scientific commentary on ‘Detection of blood‑brain barrier disruption in brains of patients with COVID‑19, but no evidence of brain penetration by SARS‑CoV‑2’,” Acta Neuropathol. 2024, 147, 1: 47. doi: https://doi.org/10.1007/s00401‑023‑02681‑y
Satta S et al., “An engineered nano‑liposome‑human ACE2 decoy neutralizes SARS‑CoV‑2 Spike protein‑induced inflammation in both murine and human macrophages,” Theranostics 2022, 12, 6: 2639–2657. doi: https://doi.org/10.7150/thno.66831
Sui Y et al., “SARS‑CoV‑2 Spike Protein Suppresses ACE2 and Type I Interferon Expression in Primary Cells From Macaque Lung Bronchoalveolar Lavage,” Front. Immunol. 2021, 12. doi: https://doi.org/10.3389/fimmu.2021.658428
Tetz G and Victor Tetz, “Prion‑Like Domains in Spike Protein of SARS‑CoV‑2 Differ across Its Variants and Enable Changes in Affinity to ACE2,” Microorganisms 2025, 10, 2: 280. doi: https://doi.org/10.3390/microorganisms10020280
Vargas‑Castro R et al., “Calcitriol prevents SARS‑CoV spike‑induced inflammation in human trophoblasts through downregulating ACE2 and TMPRSS2 expression,” J Steroid Biochem Mol Biol 2025, 245: 106625. doi: https://doi.org/10.1016/j.jsbmb.2024.106625
Youn JY et al., “Therapeutic application of estrogen for COVID‑19: Attenuation of SARS‑CoV‑2 spike protein and IL‑6 stimulated, ACE2‑dependent NOX2 activation, ROS production and MCP‑1 upregulation in endothelial cells,” Redox Biol. 2021, 46: 102099. doi: https://doi.org/10.1016/j.redox.2021.102099
Zhang S et al., “SARS‑CoV‑2 Binds Platelet ACE2 to Enhance Thrombosis in COVID‑19,” J. Hematol. Oncol. 2020, 13, 120: 120. doi: https://doi.org/10.1186/s13045‑020‑00954‑7
C. Propriétés amyloïdes, de type prion
Aksenova AY et al., “The increased amyloidogenicity of Spike RBD and pH‑dependent binding to ACE2 may contribute to the transmissibility and pathogenic properties of SARS‑CoV‑2 omicron as suggested by in silico study,” Int. J. Mol. Sci. 2022, 23, 21: 13502. doi: https://doi.org/10.3390/ijms232113502
Cao S et al., “Spike Protein Fragments Promote Alzheimer’s Amyloidogenesis,” ACS Appl. Mater. Interfaces 2023, 15, 34: 40317‑40329. doi: https://doi.org/10.1021/acsami.3c09815
Idrees D and Vijay Kumar, “SARS‑CoV‑2 Spike Protein Interactions with Amyloidogenic Proteins: Potential Clues to Neurodegeneration,” Biochemical and Biophysical Research Communications 2021, 554 : 94–98. doi: https://doi.org/10.1016/j.bbrc.2021.03.100
Ma G et al., “SARS‑CoV‑2 Spike protein S2 subunit modulates γ-secretase and enhances amyloid-β production in COVID‑19 neuropathy,” Cell Discov 2022, 8, 99. doi: https://doi.org/10.1038/s41421‑022‑00458‑3
Nahalka J, “1‑L Transcription of SARS‑CoV‑2 Spike Protein S1 Subunit,” Int. J. Mol. Sci. 2024, 25, 8: 4440. doi: https://doi.org/10.3390/ijms25084440
Petrlova J et al., “SARS‑CoV‑2 spike protein aggregation is triggered by bacterial lipopolysaccharide,” FEBS Lett. 2022, 596:2566–2575. doi: https://doi.org/10.1002/1873‑3468.14490
Petruk G et al., “SARS‑CoV‑2 spike protein binds to bacterial lipopolysaccharide and boosts proinflammatory activity,” J. Mol. Cell Biol. 2020, 12: 916‑932. doi: https://doi.org/10.1093/jmcb/mjaa067
Rong Z et al., “Persistence of spike protein at the skull‑meninges‑brain axis may contribute to the neurological sequelae of COVID‑19,” Cell Host Microbe 2024, 26: S1931‑3128(24)00438‑4. doi: https://doi.org/10.1016/j.chom.2024.11.007
Tetz G and Victor Tetz, “Prion‑Like Domains in Spike Protein of SARS‑CoV‑2 Differ across Its Variants and Enable Changes in Affinity to ACE2,” Microorganisms 2022, 10, 2: 280, doi: https://doi.org/10.3390/microorganisms10020280
Wang J et al., “SARS‑CoV‑2 Spike Protein S1 Domain Accelerates α-Synuclein Phosphorylation and Aggregation in Cellular Models of Synucleinopathy,” Mol Neurobiol. 2024, 61, 4: 2446‑2458. doi: https://doi.org/10.1007/s12035‑023‑03726‑9
D. Auto‑immune
Heil M, “Self‑DNA driven inflammation in COVID‑19 and after mRNA‑based vaccination: lessons for non‑COVID‑19 pathologies,” Front. Immunol., 2023, 14. doi: https://doi.org/10.3389/fimmu.2023.1259879
Nunez‑Castilla J et al., “Potential autoimmunity resulting from molecular mimicry between SARS- CoV‑2 spike and human proteins,” Viruses 2022, 14, 7: 1415. https://doi.org/10.3390/v14071415
E. Pression artérielle/hypertension
Angeli F et al., “The spike effect of acute respiratory syndrome coronavirus 2 and coronavirus disease 2019 vaccines on blood pressure,” Eur J Intern Med. 2023, 109: 12‑21. doi: https://doi.org/10.1016/j.ejim.2022.12.004
Sun Ǫ et al., “SARS‑coV‑2 spike protein S1 exposure increases susceptibility to angiotensin II- induced hypertension in rats by promoting central neuroinflammation and oxidative stress,” Neurochem. Res. 2023, 48, 3016–3026. doi: https://doi.org/10.1007/s11064‑023‑03949‑1
F. CD147
Avolio E et al., “The SARS‑CoV‑2 Spike Protein Disrupts Human Cardiac Pericytes Function through CD147 Receptor‑Mediated Signalling: A Potential Non‑infective Mechanism of COVID‑19 Microvascular Disease,” Clin. Sci. 2021, 135, 24: 2667–2689. doi: https://doi.org/10.1042/CS20210735
Loh D, “The potential of melatonin in the prevention and attenuation of oxidative hemolysis and myocardial injury from cd147 SARS‑CoV‑2 spike protein receptor binding,” Melatonin Research 2020, 3, 3: 380‑416. doi: https://doi.org/10.32794/mr11250069
Maugeri N et al., “Unconventional CD147‑Dependent Platelet Activation Elicited by SARS‑CoV‑2 in COVID‑19,” J. Thromb. Haemost. 2021, 20, 2: 434–448. doi: https://doi.org/10.1111/jth.15575
G. Perméabilité de la membrane cellulaire, dysfonctionnement de la barrière
Asandei A et al., “Non‑Receptor‑Mediated Lipid Membrane Permeabilization by the SARS‑CoV‑2 Spike Protein S1 Subunit,” ACS Appl. Mater. Interfaces 2020, 12, 50: 55649–55658. doi: https://doi.org/10.1021/acsami.0c17044
Biancatelli RMLC, et al. “The SARS‑CoV‑2 spike protein subunit S1 induces COVID‑19‑like acute lung injury in Kappa18‑hACE2 transgenic mice and barrier dysfunction in human endothelial cells,” Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321: L477–L484. doi: https://doi.org/10.1152/ajplung.00223.2021
Biering SB et al., “SARS‑CoV‑2 Spike Triggers Barrier Dysfunction and Vascular Leak via Integrins and TGF-β Signaling,” Nat. Commun. 2022, 13: 7630. doi: https://doi.org/10.1038/s41467‑022‑34910‑5
Buzhdygan TP et al., “The SARS‑CoV‑2 Spike Protein Alters Barrier Function in 2D Static and 3D Microfluidic in‑Vitro Models of the Human Blood‑Brain Barrier,” Neurobiol. Dis. 2020, 146: 105131. doi: https://doi.org/10.1016/j.nbd.2020.105131
Chaves JCS et al., “Differential Cytokine Responses of APOE3 and APOE4 Blood–brain Barrier Cell Types to SARS‑CoV‑2 Spike Proteins,” J. Neuroimmune Pharmacol. 2024, 19, 22. doi: https://doi.org/10.1007/s11481‑024‑10127‑9
Correa Y et al., “SARS‑CoV‑2 spike protein removes lipids from model membranes and interferes with the capacity of high‑density lipoprotein to exchange lipids,” J. Colloid Interface Sci. 2021, 602: 732- 739. doi: https://doi.org/10.1016/j.jcis.2021.06.056
DeOre BJ et al., “SARS‑CoV‑2 Spike Protein Disrupts Blood‑Brain Barrier Integrity via RhoA Activation,” J Neuroimmune Pharmacol. 2021, 16, 4:722‑728. Doi: https://doi.org/10.1007/s11481‑021‑10029‑0
Guo Y and V Kanamarlapudi, “Molecular Analysis of SARS‑CoV‑2 Spike Protein‑Induced Endothelial Cell Permeability and vWF Secretion,” Int. J. Mol. Sci. 2023, 24, 6: 5664. doi: https://doi.org/10.3390/ijms24065664
Luchini A et al., “Lipid bilayer degradation induced by SARS‑CoV‑2 spike protein as revealed by neutron reflectometry,” Sci. Rep. 2021, 11: 14867. doi: https://doi.org/10.1038/s41598‑021‑93996‑x
Luo Y et al., “SARS‑Cov‑2 spike induces intestinal barrier dysfunction through the interaction between CEACAM5 and Galectin‑9,” Front. Immunol. 2024, 15. doi: https://doi.org/10.3389/fimmu.2024.1303356
Magro N et al., “Disruption of the blood‑brain barrier is correlated with spike endocytosis by ACE2 + endothelia in the CNS microvasculature in fatal COVID‑19. Scientific commentary on ‘Detection of blood‑brain barrier disruption in brains of patients with COVID‑19, but no evidence of brain penetration by SARS‑CoV‑2’,” Acta Neuropathol. 2024, 147, 1: 47. doi: https://doi.org/10.1007/s00401‑023‑02681‑y
Raghavan S et al., “SARS‑CoV‑2 Spike Protein Induces Degradation of Junctional Proteins That Maintain Endothelial Barrier Integrity,” Front. Cardiovasc. Med. 2021, 8, 687783. doi: https://doi.org/10.3389/fcvm.2021.687783
Ruben ML et al., “The SARS‑CoV‑2 spike protein subunit S1 induces COVID‑19‑like acute lung injury in Κ18‑hACE2 transgenic mice and barrier dysfunction in human endothelial cells,” Am J Physiol Lung Cell Mol Physiol. 2021, 321, 2: L477‑L484. doi: https://doi.org/10.1152/ajplung.00223.2021
H. Cérébral, cérébrovasculaire, barrière hémato‑encéphalique, cognitif
Burnett FN et al., “SARS‑CoV‑2 Spike Protein Intensifies Cerebrovascular Complications in Diabetic hACE2 Mice through RAAS and TLR Signaling Activation,” Int. J. Mol. Sci. 2023, 24, 22: 16394.doi: https://doi.org/10.3390/ijms242216394
Choi JY et al., “SARS‑CoV‑2 spike S1 subunit protein‑mediated increase of beta‑secretase 1 (BACE1) impairs human brain vessel cells,” Biochem. Biophys. Res. Commun. 2022, 625, 20: 66‑71.doi: https://doi.org/10.1016/j.bbrc.2022.07.113
Clough E et al., “Mitochondrial Dynamics in SARS‑COV2 Spike Protein Treated Human Microglia: Implications for Neuro‑COVID,” J. Neuroimmune Pharmacol. 2021, 4, 16: 770–784. doi: https://doi.org/10.1007/s11481‑021‑10015‑6
DeOre BJ et al., “SARS‑CoV‑2 Spike Protein Disrupts Blood‑Brain Barrier Integrity via RhoA Activation,” J Neuroimmune Pharmacol. 2021, 16, 4: 722‑728. Doi: https://doi.org/10.1007/s11481‑021‑10029‑0
Erdogan MA, “Prenatal SARS‑CoV‑2 Spike Protein Exposure Induces Autism‑Like Neurobehavioral Changes in Male Neonatal Rats,” J Neuroimmune Pharmacol. 2023, 18, 4: 573‑591. doi: https://doi.org/10.1007/s11481‑023‑10089‑4
Fontes‑Dantas FL, “SARS‑CoV‑2 Spike Protein Induces TLR4‑Mediated Long- Term Cognitive Dysfunction Recapitulating Post‑COVID‑19 Syndrome in Mice,” Cell Reports 2023, 42, 3: 112189. doi: https://doi.org/10.1016/j.celrep.2023.112189
Frank MG et al., “SARS‑CoV‑2 S1 subunit produces a protracted priming of the neuroinflammatory, physiological, and behavioral responses to a remote immune challenge: A role for corticosteroids,” Brain Behav. Immun. 2024, 121: 87‑103. doi: https://doi.org/10.1016/j.bbi.2024.07.034
Heath SP et al., “SARS‑CoV‑2 Spike Protein Exacerbates Thromboembolic Cerebrovascular Complications in Humanized ACE2 Mouse Model,” Transl Stroke Res. 2024. doi: https://doi.org/10.1007/s12975‑024‑01301‑5
Khaddaj‑Mallat R et al., “SARS‑CoV‑2 deregulates the vascular and immune functions of brain pericytes via Spike protein,” Neurobiol. Dis. 2021, 161, 105561. doi: https://doi.org/10.1016/j.nbd.2021.105561
Kim ES et al., “Spike Proteins of SARS‑CoV‑2 Induce Pathological Changes in Molecular Delivery and Metabolic Function in the Brain Endothelial Cells,” Viruses 2021, 13, 10: 2021. doi: https://doi.org/10.3390/v13102021
Lykhmus O et al., “Immunization with 674–685 fragment of SARS‑Cov‑2 spike protein induces neuroinflammation and impairs episodic memory of mice,” Biochem. Biophys. Res. Commun. 2022, 622: 57–63. doi: https://doi.org/10.1016/j.bbrc.2022.07.016
Magro N et al., “Disruption of the blood‑brain barrier is correlated with spike endocytosis by ACE2 + endothelia in the CNS microvasculature in fatal COVID‑19. Scientific commentary on ‘Detection of blood‑brain barrier disruption in brains of patients with COVID‑19, but no evidence of brain penetration by SARS‑CoV‑2’,” Acta Neuropathol. 2024, 147, 1: 47. doi: https://doi.org/10.1007/s00401‑023‑02681‑y
Oh J et al., “SARS‑CoV‑2 Spike Protein Induces Cognitive Deficit and Anxiety‑Like Behavior in Mouse via Non‑cell Autonomous Hippocampal Neuronal Death,” Scientific Reports 2022, 12, 5496. doi: https://doi.org/10.1038/s41598‑022‑09410‑7
Petrovszki D et al., “Penetration of the SARS‑CoV‑2 Spike Protein across the Blood‑Brain Barrier, as Revealed by a Combination of a Human Cell Culture Model System and Optical Biosensing,” Biomedicines 2022, 10, 1: 188. doi: https://doi.org/10.3390/biomedicines10010188
Rong Z et al., “Persistence of spike protein at the skull‑meninges‑brain axis may contribute to the neurological sequelae of COVID‑19,” Cell Host Microbe 2024, 26: S1931‑3128(24)00438‑4. doi: https://doi.org/10.1016/j.chom.2024.11.007
Suprewicz L et al., “Blood‑brain barrier function in response to SARS‑CoV‑2 and its spike protein,” Neurol. Neurochir Pol. 2023, 57: 14–25. doi: https://doi.org/10.5603/PJNNS.a2023.0014
Suprewicz L et al., “Recombinant human plasma gelsolin reverses increased permeability of the blood‑brain barrier induced by the spike protein of the SARS‑CoV‑2 virus,” J Neuroinffammation 2022, 19, 1: 282, doi: https://doi.org/10.1186/s12974‑022‑02642‑4
Wu ML et al., “Mast cell activation triggered by SARS‑CoV‑2 causes inflammation in brain microvascular endothelial cells and microglia,” Front. Cell. Infect. Microbiol., 2024, 14. doi: https://doi.org/10.3389/fcimb.2024.1358873
I. Pathologie clinique
Baumeier C et al., “Intramyocardial Inflammation after COVID‑19 Vaccination: An Endomyocardial Biopsy‑Proven Case Series,” Int. J. Mol. Sci. 2022, 23: 6940. doi: https://doi.org/10.3390/ijms23136940
Craddock V et al., “Persistent circulation of soluble and extracellular vesicle‑linked Spike protein in individuals with postacute sequelae of COVID‑19,” J Med. Virol. 2023, 95, 2: e28568. doi: https://doi.org/10.1002/jmv.28568
De Michele M et al., “Evidence of SARS‑CoV‑2 Spike Protein on Retrieved Thrombi from COVID‑19 Patients,” J. Hematol. Oncol. 2022, 15, 108. doi: https://doi.org/10.1186/s13045‑022‑01329‑w
De Sousa PMB et al., “Fatal Myocarditis following COVID‑19 mRNA Immunization: A Case Report and Differential Diagnosis Review,” Vaccines 2024, 12, 2: 194.doi: https://doi.org/10.3390/vaccines12020194
Gamblicher T et al., “SARS-CoV-2 spike protein is present in both endothelial and eccrine cells of a chilblain-like skin lesion,” J Eur Acad Dermatol Venereol. 2020, 1, 10: e187‑e189. doi: https://doi.org/10.1111/jdv.16970
Gawaz A et al., “SARS‑CoV‑2–Induced Vasculitic Skin Lesions Are Associated with Massive Spike Protein Depositions in Autophagosomes,” J Invest Dermatol. 2024, 144, 2: 369‑377.e4. doi: https://doi.org/10.1016/j.jid.2023.07.018
Hulscher N et al., “Autopsy findings in cases of fatal COVID‑19 vaccine‑induced myocarditis,” ESC Heart Failure 2024. doi: https://doi.org/10.1002/ehf2.14680
Ko CJ et al., “Discordant anti‑SARS‑CoV‑2 spike protein and RNA staining in cutaneous perniotic lesions suggests endothelial deposition of cleaved spike protein,” J. Cutan Pathol 2021, 48, 1: 47–52. doi: https://doi.org/10.1111/cup.13866
Magen E et al., “Clinical and Molecular Characterization of a Rare Case of BNT162b2 mRNA COVID- 19 Vaccine‑Associated Myositis,” Vaccines 2022, 10: 1135. doi: https://doi.org/10.3390/vaccines10071135
Magro N et al., “Disruption of the blood‑brain barrier is correlated with spike endocytosis by ACE2 + endothelia in the CNS microvasculature in fatal COVID‑19. Scientific commentary on ‘Detection of blood‑brain barrier disruption in brains of patients with COVID‑19, but no evidence of brain penetration by SARS‑CoV‑2’,” Acta Neuropathol. 2024, 147, 1: 47. doi: https://doi.org/10.1007/s00401‑023‑02681‑y
Mörz M, “A Case Report: Multifocal Necrotizing Encephalitis and Myocarditis after BNT162b2 mRNA Vaccination against COVID‑19,” Vaccines 2022, 10, 10: 1651. doi: https://doi.org/10.3390/vaccines10101651
Rong Z et al., “Persistence of spike protein at the skull‑meninges‑brain axis may contribute to the neurological sequelae of COVID‑19,” Cell Host Microbe 2024, 26: S1931‑3128(24)00438‑4. doi: https://doi.org/10.1016/j.chom.2024.11.007
Sano H et al., “A case of persistent, confluent maculopapular erythema following a COVID‑19 mRNA vaccination is possibly associated with the intralesional spike protein expressed by vascular endothelial cells and eccrine glands in the deep dermis,” J. Dermatol. 2023, 50: 1208–1212. doi: https://doi.org/10.1111/1346‑8138.16816
Sano S et al., “SARS‑CoV‑2 spike protein found in the acrosyringium and eccrine gland of repetitive miliaria‑like lesions in a woman following mRNA vaccination,” J. Dermatol. 2024, 51, 9: e293‑e295. doi: https://doi.org/10.1111/1346‑8138.17204
Santonja C et al., “COVID‑19 chilblain‑like lesion: immunohistochemical demonstration of SARS- CoV‑2 spike protein in blood vessel endothelium and sweat gland epithelium in a polymerase chain reaction‑negative patient,” Br J Dermatol. 2020, 183, 4: 778‑780. doi: https://doi.org/10.1111/bjd.19338
Soares CD et al., “Oral vesiculobullous lesions as an early sign of COVID-19: immunohistochemical detection of SARS-CoV-2 spike protein,” Br. J. Dermatol. 2021, 184, 1: e6.doi: https://doi.org/10.1111/bjd.19569
Yamamoto M et al., “Persistent varicella zoster virus infection following mRNA COVID‑19 vaccination was associated with the presence of encoded spike protein in the lesion,” J. Cutan Immunol. Allergy. 2022: 1‑6. doi: https://doi.org/10.1002/cia2.12278
Al‑Kuraishy HM et al., “Changes in the Blood Viscosity in Patients With SARS‑CoV‑2 Infection,” Front. Med. 2022, 9: 876017. doi: https://doi.org/10.3389/fmed.2022.876017
Appelbaum K et al., “SARS‑CoV‑2 spike‑dependent platelet activation in COVID‑19 vaccine‑induced thrombocytopenia,” Blood Adv. 2022, 6: 2250–2253. doi: https://doi.org/10.1182/bloodadvances.2021005050
Boschi C et al., “SARS‑CoV‑2 Spike Protein Induces Hemagglutination: Implications for COVID‑19 Morbidities and Therapeutics and for Vaccine Adverse Effects,” Int. J. Biol. Macromol. 2022, 23, 24: 15480, doi: https://doi.org/10.3390/ijms232415480
Bye AP et al., “Aberrant glycosylation of anti‑SARS‑CoV‑2 spike IgG is a prothrombotic stimulus for platelets,” Blood 2021, 138, 6: 1481–9. doi: https://doi.org/10.1182/blood.2021011871
Carnevale R et al., “Toll‑Like Receptor 4‑Dependent Platelet‑Related Thrombosis in SARS‑CoV‑2 Infection,” Circ. Res. 2023, 132, 3: 290– 305, doi: https://doi.org/10.1161/CIRCRESAHA.122.321541
Cossenza LC et al., “Inhibitory effects of SARS‑CoV‑2 spike protein and BNT162b2 vaccine on erythropoietin‑induced globin gene expression in erythroid precursor cells from patients with β- thalassemia,” Exp. Hematol. 2024, 129, 104128. doi: https://doi.org/10.1016/j.exphem.2023.11.002
De Michele M et al., “Vaccine‑induced immune thrombotic thrombocytopenia: a possible pathogenic role of ChAdOx1 nCoV‑19 vaccine‑encoded soluble SARS‑CoV‑2 spike protein,” Haematologica 2022, 107, 7: 1687–92. https://doi.org/10.3324/haematol.2021.280180
Grobbelaar LM et al., “SARS‑CoV‑2 Spike Protein S1 Induces Fibrin(ogen) Resistant to Fibrinolysis: Implications for Microclot Formation in COVID‑19,” Biosicence Reports 2021, 41, 8: BSR20210611. doi: https://doi.org/10.1042/BSR20210611
Heath SP et al., “SARS‑CoV‑2 Spike Protein Exacerbates Thromboembolic Cerebrovascular Complications in Humanized ACE2 Mouse Model,” Transl Stroke Res. 2024. doi: https://doi.org/10.1007/s12975‑024‑01301‑5
Iba T and JH Levy, “The roles of platelets in COVID‑19‑associated coagulopathy and vaccine‑induced immune thrombotic thrombocytopenia,” Trends Cardiovasc Med. 2022, 32, 1: 1‑9. doi: https://doi.org/10.1016/j.tcm.2021.08.012
Huynh TV et al., “Spike Protein of SARS‑CoV‑2 Activates Cardiac Fibrogenesis through NLRP3 Inflammasomes and NF-κB Signaling,” Cells 2024, 13, 16: 1331.doi: https://doi.org/10.3390/cells13161331
Iba T and JH Levy, “The roles of platelets in COVID‑19‑associated coagulopathy and vaccine‑induced immune thrombotic thrombocytopenia,” Trends Cardiovasc Med. 2022, 32, 1: 1‑9. doi: https://doi.org/10.1016/j.tcm.2021.08.012
Jana S et al., “Cell‑free hemoglobin does not attenuate the effects of SARS‑CoV‑2 spike protein S1 subunit in pulmonary endothelial cells,” Int. J. Mol. Sci. 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Kim SY et al., “Characterization of heparin and severe acute respiratory syndrome‑related coronavirus 2 (SARS‑CoV‑2) spike glycoprotein binding interactions,” Antivir Res. 2020, 181: 104873. doi: https://doi.org/10.1016/j.antiviral.2020.104873
Kircheis R, “Coagulopathies after Vaccination against SARS‑CoV‑2 May Be Derived from a Combined Effect of SARS‑CoV‑2 Spike Protein and Adenovirus Vector‑Triggered Signaling Pathways,” Int. J. Mol. Sci. 2021, 22, 19: 10791. https://doi.org/10.3390/ijms221910791
Kuhn CC et al. “Direct Cryo‑ET observation of platelet deformation induced by SARS‑CoV‑2 spike protein,” Nat. Commun. 2023, 14, 620. doi: https://doi.org/10.1038/s41467‑023‑36279‑5
Li T et al., “Platelets Mediate Inflammatory Monocyte Activation by SARS‑CoV‑2 Spike Protein,” J. Clin. Invest. 2022, 132, 4: e150101. doi: https://doi.org/10.1172/JCI150101
Maugeri N et al., “Unconventional CD147‑Dependent Platelet Activation Elicited by SARS‑CoV‑2 in COVID‑19,” J. Thromb. Haemost. 2021, 20, 2: 434–448, doi: https://doi.org/10.1111/jth.15575
Passariello M et al., “Interactions of Spike‑RBD of SARS‑CoV‑2 and Platelet Factor 4: New Insights in the Etiopathogenesis of Thrombosis,” Int. J. Mol. Sci. 2021, 22, 16: 8562.doi: https://doi.org/10.3390/ijms22168562
Perico L et al., “SARS‑CoV‑2 Spike Protein 1 Activates Microvascular Endothelial Cells and Complement System Leading to Platelet Aggregation,” Front. Immunol. 2022, 13, 827146. doi: https://doi.org/10.3389/fimmu.2022.827146
Roytenberg R et al., “Thymidine phosphorylase mediates SARS‑CoV‑2 spike protein enhanced thrombosis in K18‑hACE2TG mice,” Thromb. Res. 2024, 244, 8: 109195. doi: https://doi.org/10.1016/j.thromres.2024.109195
Russo A, et al., “Implication of COVID‑19 on Erythrocytes Functionality: Red Blood Cell Biochemical Implications and Morpho‑Functional Aspects,” Int. J. Mol. Sci. 2022, 23, 4: 2171.doi: https://doi.org/10.3390/ijms23042171
Scheim, DE. “A Deadly Embrace: Hemagglutination Mediated by SARS‑CoV‑2 Spike Protein at its 22 N‑Glycosylation Sites, Red Blood Cell Surface Sialoglycoproteins, and Antibody,” Int. J. Mol.Sci. 2022, 23, 5: 2558. doi: https://doi.org/10.3390/ijms23052558
Scheim DE et al., “Sialylated Glycan Bindings from SARS‑CoV‑2 Spike Protein to Blood and Endothelial Cells Govern the Severe Morbidities of COVID‑19,” Int. J. Mol. Sci. 2023, 24, 23: 17039. doi: https://doi.org/10.3390/ijms242317039
Sirsendu J et al., “Cell‑Free Hemoglobin Does Not Attenuate the Effects of SARS‑CoV‑2 Spike Protein S1 Subunit in Pulmonary Endothelial Cells,” Int. J. Mol. Sci., 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Zhang S et al., “SARS‑CoV‑2 Binds Platelet ACE2 to Enhance Thrombosis in COVID‑19,” J. Hematol. Oncol. 2020, 13, 120: 120. doi: https://doi.org/10.1186/s13045‑020‑00954‑7
Zhang Z et al., “SARS‑CoV‑2 spike protein dictates syncytium‑mediated lymphocyte elimination,” Cell Death Differ. 2021, 28, 2765–2777. doi: https://doi.org/10.1038/s41418‑021‑00782‑3
Zheng Y et al., “SARS‑CoV‑2 Spike Protein Causes Blood Coagulation and Thrombosis by Competitive Binding to Heparan Sulfate,” Int. J. Biol. Macromol. 2021, 193: 1124–1129. doi: https://doi.org/10.1016/j.ijbiomac.2021.10.112
K. Cytokines, chemokines, interféron, interleukins
Ao Z et al., “SARS‑CoV‑2 Delta spike protein enhances the viral fusogenicity and inflammatory cytokine production,” iScience 2022, 25, 8: 104759. doi: https://doi.org/10.1016/j.isci.2022.104759
Chaves JCS et al., “Differential Cytokine Responses of APOE3 and APOE4 Blood–brain Barrier Cell Types to SARS‑CoV‑2 Spike Proteins,” J. Neuroimmune Pharmacol. 2024, 19, 22. doi: https://doi.org/10.1007/s11481‑024‑10127‑9
Chittasupho C et al., “Targeting spike glycoprotein S1 mediated by NLRP3 inflammasome machinery and the cytokine releases in A549 lung epithelial cells by nanocurcumin,” Pharmaceuticals (Basel) 2023, 16, 6: 862. doi: https://doi.org/10.3390/ph16060862
Das T et al., “N‑glycosylation of the SARS‑CoV‑2 spike protein at Asn331 and Asn343 is involved in spike‑ACE2 binding, virus entry, and regulation of IL‑6,” Microbiol. Immunol. 2024, 68, 5: 165‑178. doi: https://doi.org/10.1111/1348‑0421.13121
Duarte C, “Age‑dependent effects of the recombinant spike protein/SARS‑CoV‑2 on the M‑CSF- and IL‑34‑differentiated macrophages in vitro,” Biochem. Biophys. Res. Commun. 2021, 546: 97–102. doi: https://doi.org/10.1016/j.bbrc.2021.01.104
Forsyth CB et al., “The SARS‑CoV‑2 S1 spike protein promotes MAPK and NF‑kB activation in human lung cells and inflammatory cytokine production in human lung and intestinal epithelial cells,” Microorganisms 2022, 10, 10: 1996. doi: https://doi.org/10.3390/microorganisms10101996
Freitas RS et al., “SARS‑CoV‑2 Spike antagonizes innate antiviral immunity by targeting interferon regulatory factor 3,” Front Cell Infect Microbiol. 2021, 11: 789462. doi: https://doi.org/10.3389/fcimb.2021.789462
Gasparello J et al., “Sulforaphane inhibits the expression of interleukin‑6 and interleukin‑8 induced in bronchial epithelial IB3‑1 cells by exposure to the SARS‑CoV‑2 Spike protein,” Phytomedicine 2021, 87: 153583. doi: https://doi.org/10.1016/j.phymed.2021.153583
Ghazanfari D et al., “Mechanistic insights into SARS‑CoV‑2 spike protein induction of the chemokine CXCL10,” Sci. Rep. 2024, 14: 11179. doi: https://doi.org/10.1038/s41598‑024‑61906‑6
Gracie NP et al., “Cellular signalling by SARS‑CoV‑2 spike protein,” Microbiology Australia 2024, 45, 1: 13‑17. doi: https://doi.org/10.1071/MA24005
Gu T et al., “Cytokine Signature Induced by SARS‑CoV‑2 Spike Protein in a Mouse Model,” Front. Immunol., 2021 (Sec. Inflammation). doi: https://doi.org/10.3389/fimmu.2020.621441
Jugler C et al, “SARS‑CoV‑2 Spike Protein‑Induced Interleukin 6 Signaling Is Blocked by a Plant- Produced Anti‑Interleukin 6 Receptor Monoclonal Antibody,” Vaccines 2021, S, 11 1365. https://doi.org/10.3390/vaccines9111365
Liang S et al., “SARS‑CoV‑2 spike protein induces IL‑18‑mediated cardiopulmonary inflammation via reduced mitophagy,” Signal Transduct Target Ther 2023, 8, 103. doi: https://doi.org/10.1038/s41392‑023‑01368‑w
Liu T et al., “RS‑5645 attenuates inflammatory cytokine storm induced by SARS‑CoV‑2 spike protein and LPS by modulating pulmonary microbiota,” Int. J. Biol. Sci. 2021, 17, 13: 3305–3319. doi: https://doi.org/10.7150/ijbs.63329
Liu X et al., “SARS‑CoV‑2 spike protein‑induced cell fusion activates the cGAS‑STING pathway and the interferon response,” Sci Signal. 2022, 15, 729: eabg8744. doi: https://doi.org/10.1126/scisignal.abg8744
Niu C et al., “SARS‑CoV‑2 spike protein induces the cytokine release syndrome by stimulating T cells to produce more IL‑2,” Front. Immunol. 2024, 15: 1444643. doi: https://doi.org/10.3389/fimmu.2024.1444643
Norris B et al., “Evaluation of Glutathione in Spike Protein of SARS‑CoV‑2 Induced Immunothrombosis and Cytokine Dysregulation,” Antioxidants 2024, 13, 3: 271. doi: https://doi.org/10.3390/antiox13030271
Olajide OA et al., “Induction of Exaggerated Cytokine Production in Human Peripheral Blood Mononuclear Cells by a Recombinant SARS‑CoV‑2 Spike Glycoprotein S1 and Its Inhibition by Dexamethasone,” Inffammation 2021, 44: 1865–1877. doi: https://doi.org/10.1007/s10753‑021‑01464‑5
Park YJ et al., “D‑dimer and CoV‑2 spike‑immune complexes contribute to the production of PGE2 and proinflammatory cytokines in monocytes,” PLoS Pathog., 2022, 18, 4: e1010468. doi: https://doi.org/10.1371/journal.ppat.1010468
Patra T et al., “SARS‑CoV‑2 spike protein promotes IL‑6 trans‑signaling by activation of angiotensin II receptor signaling in epithelial cells,” PLoS Pathog. 2020, 16: e1009128. doi: https://doi.org/10.1371/journal.ppat.1009128
Samsudin S et al., “SARS‑CoV‑2 spike protein as a bacterial lipopolysaccharide delivery system in an overzealous inflammatory cascade,” J. Mol. Biol. 2022, 14, 9: mjac058. doi: https://doi.org/10.1093/jmcb/mjac058
Schroeder JT and AP Bieneman, “The S1 Subunit of the SARS‑CoV‑2 Spike protein activates human monocytes to produce cytokines linked to COVID‑19: relevance to galectin‑3,” Front Immunol. 2022, 13: 831763. doi: https://doi.org/10.3389/fimmu.2022.831763
Sharma VK et al., “Nanocurcumin Potently Inhibits SARS‑CoV‑2 Spike Protein‑Induced Cytokine Storm by Deactivation of MAPK/NF-κB Signaling in Epithelial Cells,” ACS Appl. Bio Mater. 2022, 5, 2: 483–491. doi: https://doi.org/10.1021/acsabm.1c00874
Sui Y et al., “SARS‑CoV‑2 Spike Protein Suppresses ACE2 and Type I Interferon Expression in Primary Cells From Macaque Lung Bronchoalveolar Lavage,” Front. Immunol. 2021, 12. doi: https://doi.org/10.3389/fimmu.2021.658428
Youn JY et al., “Therapeutic application of estrogen for COVID‑19: Attenuation of SARS‑CoV‑2 spike protein and IL‑6 stimulated, ACE2‑dependent NOX2 activation, ROS production and MCP‑1 upregulation in endothelial cells,” Redox Biol. 2021, 46: 102099. doi: https://doi.org/10.1016/j.redox.2021.102099
Zhang Ǫ et al., “Severe acute respiratory syndrome coronavirus 2 (SARS‑CoV‑2) membrane (M) and spike (S) proteins antagonize host type i interferon response,” Front Cell Infect Microbiol 2021, 11: 766922. doi: https://doi.org/10.3389/fcimb.2021.766922
Zhang RG et al., “SARS‑CoV‑2 spike protein receptor binding domain promotes IL‑6 and IL‑8 release via ATP/P2Y2 and ERK1/2 signaling pathways in human bronchial epithelia,” Mol. Immunol. 2024, 167: 53‑61. doi: https://doi.org/10.1016/j.molimm.2024.02.005
L. Endothélial
Bhargavan B and GD Kanmogne, “SARS‑CoV‑2 spike proteins and cell–cell communication inhibits TFPI and induces thrombogenic factors in human lung microvascular endothelial cells and neutrophils: implications for COVID‑19 coagulopathy pathogenesis,” Int. J. Mol. Sci. 2022, 23, 18: 10436. doi: https://doi.org/10.3390/ijms231810436
Biancatelli RMLC, et al. “The SARS‑CoV‑2 spike protein subunit S1 induces COVID‑19‑like acute lung injury in Kappa18‑hACE2 transgenic mice and barrier dysfunction in human endothelial cells,” Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321: L477–L484. doi: https://doi.org/10.1152/ajplung.00223.2021
Gamblicher T et al., “SARS-CoV-2 spike protein is present in both endothelial and eccrine cells of a chilblain-like skin lesion,” J Eur Acad Dermatol Venereol. 2020, 1, 10: e187‑e189. doi: https://doi.org/10.1111/jdv.16970
Guo Y and V Kanamarlapudi, “Molecular Analysis of SARS‑CoV‑2 Spike Protein‑Induced Endothelial Cell Permeability and vWF Secretion,” Int. J. Mol. Sci. 2023, 24, 6: 5664.doi: https://doi.org/10.3390/ijms24065664
Jana S et al., “Cell‑free hemoglobin does not attenuate the effects of SARS‑CoV‑2 spike protein S1 subunit in pulmonary endothelial cells,” Int. J. Mol. Sci. 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Kulkoviene G et al., “Differential Mitochondrial, Oxidative Stress and Inflammatory Responses to SARS‑CoV‑2 Spike Protein Receptor Binding Domain in Human Lung Microvascular, Coronary Artery Endothelial and Bronchial Epithelial Cells,” Int. J. Mol. Sci. 2024, 25, 6: 3188.doi: https://doi.org/10.3390/ijms25063188
Marrone L et al., “Tirofiban prevents the effects of SARS‑CoV‑2 spike protein on macrophage activation and endothelial cell death,” Heliyon, 2024, 10, 15, e35341. doi: https://doi.org/10.1016/j.heliyon.2024.e35341
Meyer K et al., “SARS‑CoV‑2 Spike Protein Induces Paracrine Senescence and Leukocyte Adhesion in Endothelial Cells,” J. Virol. 2021, 95: e0079421. doi: https://doi.org/10.1128/jvi.00794‑21
Nuovo JG et al., “Endothelial Cell Damage Is the Central Part of COVID‑19 and a Mouse Model Induced by Injection of the S1 Subunit of the Spike Protein,” Ann. Diagn. Pathol. 2021, 51, 151682. doi: https://doi.org/10.1016/j.anndiagpath.2020.151682
Perico L et al., “SARS‑CoV‑2 Spike Protein 1 Activates Microvascular Endothelial Cells and Complement System Leading to Platelet Aggregation,” Front. Immunol. 2022, 13, 827146. doi: https://doi.org/10.3389/fimmu.2022.827146
Raghavan S et al., “SARS‑CoV‑2 Spike Protein Induces Degradation of Junctional Proteins That Maintain Endothelial Barrier Integrity,” Front. Cardiovasc. Med. 2021, 8, 687783. doi: https://doi.org/10.3389/fcvm.2021.687783
Ratajczak MZ et al., “SARS‑CoV‑2 Entry Receptor ACE2 Is Expressed on Very Small CD45- Precursors of Hematopoietic and Endothelial Cells and in Response to Virus Spike Protein Activates the Nlrp3 Inflammasome,” Stem Cell Rev Rep. 2021, 17, 1: 266‑277. doi: https://doi.org/10.1007/s12015‑020- 10010‑z
Robles JP et al., “The Spike Protein of SARS‑CoV‑2 Induces Endothelial Inflammation through Integrin α5β1 and NF-κB Signaling,” J. Biol. Chem. 2022, 298, 3: 101695. doi: https://doi.org/10.1016/j.jbc.2022.101695
Rotoli BM et al., “Endothelial cell activation by SARS‑CoV‑2 spike S1 protein: A crosstalk between endothelium and innate immune cells,” Biomedicines 2021, 9, 9: 1220. doi: https://doi.org/10.3390/biomedicines9091220
Ruben ML et al., “The SARS‑CoV‑2 spike protein subunit S1 induces COVID‑19‑like acute lung injury in Κ18‑hACE2 transgenic mice and barrier dysfunction in human endothelial cells,” Am J Physiol Lung Cell Mol Physiol. 2021, 321, 2: L477‑L484. doi: https://doi.org/10.1152/ajplung.00223.2021
Sano H et al., “A case of persistent, confluent maculopapular erythema following a COVID‑19 mRNA vaccination is possibly associated with the intralesional spike protein expressed by vascular endothelial cells and eccrine glands in the deep dermis,” J. Dermatol. 2023, 50: 1208–1212. doi: https://doi.org/10.1111/1346‑8138.16816
Santonja C et al., “COVID‑19 chilblain‑like lesion: immunohistochemical demonstration of SARS- CoV‑2 spike protein in blood vessel endothelium and sweat gland epithelium in a polymerase chain reaction‑negative patient,” Br J Dermatol. 2020, 183, 4: 778‑780. doi: https://doi.org/10.1111/bjd.19338
Scheim DE et al., “Sialylated Glycan Bindings from SARS‑CoV‑2 Spike Protein to Blood and Endothelial Cells Govern the Severe Morbidities of COVID‑19,” Int. J. Mol. Sci. 2023, 24, 23: 17039. doi: https://doi.org/10.3390/ijms242317039
Sirsendu J et al., “Cell‑Free Hemoglobin Does Not Attenuate the Effects of SARS‑CoV‑2 Spike Protein S1 Subunit in Pulmonary Endothelial Cells,” Int. J. Mol. Sci., 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Stern B et al., “SARS‑CoV‑2 spike protein induces endothelial dysfunction in 3D engineered vascular networks,” J. Biomed. Mater. Res. A. 2023, 112, 4: 524‑533. doi: https://doi.org/10.1002/jbm.a.37543
Villacampa A et al., “SARS‑CoV‑2 S protein activates NLRP3 inflammasome and deregulates coagulation factors in endothelial and immune cells,” Cell Commun. Signal. 2024, 22, 38. doi: https://doi.org/10.1186/s12964‑023‑01397‑6
Wu ML et al., “Mast cell activation triggered by SARS‑CoV‑2 causes inflammation in brain microvascular endothelial cells and microglia,” Front. Cell. Infect. Microbiol. 2024, 14. doi: https://doi.org/10.3389/fcimb.2024.1358873
Youn JY et al., “Therapeutic application of estrogen for COVID‑19: Attenuation of SARS‑CoV‑2 spike protein and IL‑6 stimulated, ACE2‑dependent NOX2 activation, ROS production and MCP‑1 upregulation in endothelial cells,” Redox Biol. 2021, 46: 102099. doi: https://doi.org/10.1016/j.redox.2021.102099
Zekri‑Nechar K et al., “Spike Protein Subunits of SARS‑CoV‑2 Alter Mitochondrial Metabolism in Human Pulmonary Microvascular Endothelial Cells: Involvement of Factor Xa,” Dis. Markers 2022, 1118195. doi: https://doi.org/10.1155/2022/1118195
M. Gastro‑intestinal
Forsyth CB et al., “The SARS‑CoV‑2 S1 spike protein promotes MAPK and NF‑kB activation in human lung cells and inflammatory cytokine production in human lung and intestinal epithelial cells,” Microorganisms 2022, 10, 10: 1996. doi: https://doi.org/10.3390/microorganisms10101996
Li Z et al., “SARS‑CoV‑2 pathogenesis in the gastrointestinal tract mediated by Spike‑induced intestinal inflammation,” Precis. Clin. Med. 2024, 7, 1: pbad034. doi: https://doi.org/10.1093/pcmedi/pbad034
Luo Y et al., “SARS‑Cov‑2 spike induces intestinal barrier dysfunction through the interaction between CEACAM5 and Galectin‑9,” Front. Immunol. 2024, 15. doi: https://doi.org/10.3389/fimmu.2024.1303356
Nascimento RR et al., “SARS‑CoV‑2 Spike protein triggers gut impairment since mucosal barrier to innermost layers: From basic science to clinical relevance,” Mucosal Immunol. 2024, 17, 4: 565‑583. doi: https://doi.org/10.1016/j.mucimm.2024.03.00
Yilmaz A et al., “Differential proinflammatory responses of colon epithelial cells to SARS‑CoV‑2 spike protein and Pseudomonas aeruginosa lipopolysaccharide,” Turk J Biochem. 2024. doi: https://doi.org/10.1515/tjb‑2024‑0144
Zeng FM et al., “SARS‑CoV‑2 spike spurs intestinal inflammation via VEGF production in enterocytes,” EMBO Mol Med. 2022, 14: e14844. doi: https://doi.org/10.15252/emmm.202114844
N. Dysfonctionnement immunitaire
Baldari CT et al., “Emerging Roles of SARS‑CoV‑2 Spike‑ACE2 in Immune Evasion and Pathogenesis,” Trends Immunol. 2023, 44, 6. doi: https://doi.org/10.1016/j.it.2023.04.001
Bocquet‑Garcon A, “Impact of the SARS‑CoV‑2 Spike Protein on the Innate Immune System: A Review,” Cureus 2024, 16, 3: e57008. doi: https://doi.org/10.7759/cureus.57008
Kim MJ et al., “The SARS-CoV-2 spike protein induces lung cancer migration and invasion in a TLR2- dependent manner,” Cancer Commun (London), 2023, 44, 2: 273–277.doi: https://doi.org/10.1002/cac2.12485
Onnis A et al., “SARS‑CoV‑2 Spike protein suppresses CTL‑mediated killing by inhibiting immune synapse assembly,” J Exp Med 2023, 220, 2: e20220906. doi: https://doi.org/10.1084/jem.20220906
O. Macrophages, monocytes, neutrophiles
Ahn WM et al., “SARS‑CoV‑2 Spike Protein Stimulates Macropinocytosis in Murine and Human Macrophages via PKC‑NADPH Oxidase Signaling,” Antioxidants 2024, 13, 2: 175.doi: https://doi.org/10.3390/antiox13020175
Ait‑Belkacem I et al., “SARS‑CoV‑2 spike protein induces a differential monocyte activation that may contribute to age bias in COVID‑19 severity,” Sci. Rep. 2022, 12: 20824. doi: ttps://doi.org/10.1038/s41598‑022‑25259‑2
Barhoumi T et al., “SARS‑CoV‑2 coronavirus Spike protein‑induced apoptosis, inflammatory, and oxidative stress responses in THP‑1-like‑macrophages: potential role of angiotensin‑converting enzyme inhibitor (perindopril),” Front. Immunol. 2021, 12: 728896. doi: https://doi.org/10.3389/fimmu.2021.728896
Bortolotti D et al., “SARS‑CoV‑2 Spike 1 Protein Controls Natural Killer Cell Activation via the HLA- E/NKG2A Pathway,” Cells 2020, 9, 9: 1975. doi: https://doi.org/10.3390/cells9091975
Cao X et al., “Spike protein of SARS‑CoV‑2 activates macrophages and contributes to induction of acute lung inflammation in male mice,” FASEB J. 2021, 35, e21801. doi: https://doi.org/10.1096/fj.202002742RR
Chiok K et al., “Proinflammatory Responses in SARS‑CoV‑2 and Soluble Spike Glycoprotein S1 Subunit Activated Human Macrophages,” Viruses 2023, 15, 3: 754.doi: https://doi.org/10.3390/v15030754
Cory TJ et al., “Metformin Suppresses Monocyte Immunometabolic Activation by SARS‑CoV‑2 Spike Protein Subunit 1,” Front. Immunol. 2021, 12 (Sec. Cytokines and Soluble Mediators in Immunity): 733921. doi: https://doi.org/10.3389/fimmu.2021.733921
Del Re A et al., “Ultramicronized Palmitoylethanolamide Inhibits NLRP3 Inflammasome Expression and Pro‑Inflammatory Response Activated by SARS‑CoV‑2 Spike Protein in Cultured Murine Alveolar Macrophages,” Metabolites 2021, 11, 9: 592. doi: https://doi.org/10.3390/metabo11090592
Duarte C, “Age‑dependent effects of the recombinant spike protein/SARS‑CoV‑2 on the M‑CSF- and IL‑34‑differentiated macrophages in vitro,” Biochem. Biophys. Res. Commun. 2021, 546: 97–102. doi: https://doi.org/10.1016/j.bbrc.2021.01.104
Karwaciak I et al., “Nucleocapsid and Spike Proteins of the Coronavirus Sars‑Cov‑2 Induce Il6 in Monocytes and Macrophages—Potential Implications for Cytokine Storm Syndrome,” Vaccines 2021, 9, 1, 54: 1–10. doi: https://doi.org/10.3390/vaccines9010054
Li T et al., “Platelets Mediate Inflammatory Monocyte Activation by SARS‑CoV‑2 Spike Protein,” J. Clin. Invest. 2022, 132, 4: e150101. doi: https://doi.org/10.1172/JCI150101
Loh JT et al., “Dok3 restrains neutrophil production of calprotectin during TLR4 sensing of SARS‑CoV- 2 spike protein,” Front. Immunol. 2022, 13 (Sec. Molecular Innate Immunity). doi: https://doi.org/10.3389/fimmu.2022.996637
Marrone L et al., “Tirofiban prevents the effects of SARS‑CoV‑2 spike protein on macrophage activation and endothelial cell death,” Heliyon, 2024, 10, 15: e35341. doi: https://doi.org/10.1016/j.heliyon.2024.e35341
Miller GM et al., “SARS‑CoV‑2 and SARS‑CoV‑2 Spike protein S1 subunit Trigger Proinflammatory Response in Macrophages in the Absence of Productive Infection,” J. Immunol. 2023, 210 (1_Supplement): 71.30. doi: https://doi.org/10.4049/jimmunol.210.Supp.71.30
Onnis A et al., “SARS‑CoV‑2 Spike protein suppresses CTL‑mediated killing by inhibiting immune synapse assembly,” J Exp Med 2023, 220, 2: e20220906. doi: https://doi.org/10.1084/jem.20220906
Palestra F et al. “SARS‑CoV‑2 Spike Protein Activates Human Lung Macrophages,” Int. J. Mol. Sci. 2023, 24, 3: 3036. doi: https://doi.org/10.3390/ijms24033036
Park C et al., “Murine alveolar Macrophages Rapidly Accumulate intranasally Administered SARS- CoV‑2 Spike Protein leading to neutrophil Recruitment and Damage,” Elife 2024, 12, RP86764. doi: https://doi.org/10.7554/eLife.86764.3
Park YJ et al., “D‑dimer and CoV‑2 spike‑immune complexes contribute to the production of PGE2 and proinflammatory cytokines in monocytes,” PLoS Pathog. 2022, 18, 4: e1010468. doi: https://doi.org/10.1371/journal.ppat.1010468
Park YJ et al., “Pyrogenic and inflammatory mediators are produced by polarized M1 and M2 macrophages activated with D‑dimer and SARS‑CoV‑2 spike immune complexes,” Cytokine 2024, 173: 156447. doi: https://doi.org/10.1016/j.cyto.2023.156447
Patterson BK et al., “Persistence of SARS CoV‑2 S1 Protein in CD16+ Monocytes in Post‑Acute Sequelae of COVID‑19 (PASC) up to 15 Months Post‑Infection,” Front. Immunol. 12 (Sec. Viral Immunology). doi: https://doi.org/10.3389/fimmu.2021.746021
Pence B, “Recombinant SARS‑CoV‑2 Spike Protein Mediates Glycolytic and Inflammatory Activation in Human Monocytes,” Innov Aging 2020, 4, sp. 1: 955. doi: https://doi.org/10.1093/geroni/igaa057.3493
Satta S et al., “An engineered nano‑liposome‑human ACE2 decoy neutralizes SARS‑CoV‑2 Spike protein‑induced inflammation in both murine and human macrophages,” Theranostics 2022, 12, 6: 2639–2657. doi: https://doi.org/10.7150/thno.66831
Schroeder JT and AP Bieneman, “The S1 Subunit of the SARS‑CoV‑2 Spike protein activates human monocytes to produce cytokines linked to COVID‑19: relevance to galectin‑3,” Front Immunol. 2022, 13: 831763. doi: https://doi.org/10.3389/fimmu.2022.831763
Shirato K and Takako Kizaki, “SARS‑CoV‑2 Spike Protein S1 Subunit Induces Pro- inflammatory Responses via Toll‑Like Receptor 4 Signaling in Murine and Human Macrophages,” Heliyon 2021, 7, 2: e06187, doi: https://doi.org/10.1016/j.heliyon.2021.e06187
Vettori M et al., “Effects of Different Types of Recombinant SARS‑CoV‑2 Spike Protein on Circulating Monocytes’ Structure,” Int. J. Mol. Sci. 2023, 24, 11, 9373. doi: https://doi.org/10.3390/ijms24119373
Youn YJ et al., “Nucleocapsid and spike proteins of SARS‑CoV‑2 drive neutrophil extracellular trap formation,” Immune Netw. 2021, 21, 2: e16. doi: https://doi.org/10.4110/in.2021.21.e16
Zaki H and S Khan, “SARS‑CoV‑2 spike protein induces inflammatory molecules through TLR2 in macrophages and monocytes,” J. Immunol. 2021, 206 (1_supplement): 62.07. doi: https://doi.org/10.4049/jimmunol.206.Supp.62.07
P. MAPK/NF‑kB
Arjsri P et al., “Hesperetin from root extract of Clerodendrum petasites S. Moore inhibits SARS‑CoV‑2 spike protein S1 subunit‑induced Nlrp3 inflammasome in A549 lung cells via modulation of the Akt/Mapk/Ap‑1 pathway,” Int. J. Mol. Sci. 2022, 23, 18: 10346. doi: https://doi.org/10.3390/ijms231810346
Bhattacharyya S and JK Tobacman, “SARS‑CoV‑2 spike protein‑ACE2 interaction increases carbohydrate sulfotransferases and reduces N‑acetylgalactosamine‑4-sulfatase by p38 MAPK,” Signal Transduct Target Ther 2024, 9, 39. doi: https://doi.org/10.1038/s41392‑024‑01741‑3
Forsyth CB et al., “The SARS‑CoV‑2 S1 spike protein promotes MAPK and NF‑kB activation in human lung cells and inflammatory cytokine production in human lung and intestinal epithelial cells,” Microorganisms 2022, 10, 10: 1996. doi: https://doi.org/10.3390/microorganisms10101996
Johnson EL et al., “The S1 spike protein of SARS‑CoV‑2 upregulates the ERK/MAPK signaling pathway in DC‑SIGN‑expressing THP‑1 cells,” Cell Stress Chaperones 2024, 29, 2: 227‑234. doi: https://doi.org/10.1016/j.cstres.2024.03.002
Khan S et al., “SARS‑CoV‑2 Spike Protein Induces Inflammation via TLR2‑Dependent Activation of the NF-κB Pathway,” eLife 2021, 10: e68563, doi: https://doi.org/10.7554/elife.68563
Kircheis R and O Planz, “Could a Lower Toll‑like Receptor (TLR) and NF-κB Activation Due to a Changed Charge Distribution in the Spike Protein Be the Reason for the Lower Pathogenicity of Omicron?” Int. J. Mol. Sci. 2022, 23, 11: 5966. doi: https://doi.org/10.3390/ijms23115966
Kyriakopoulos AM et al., “Mitogen Activated Protein Kinase (MAPK) Activation, p53, and Autophagy Inhibition Characterize the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS‑CoV‑2) Spike Protein Induced Neurotoxicity,” Cureus 2022, 14, 12: e32361. doi: https://doi.org/10.7759/cureus.32361
Robles JP et al., “The Spike Protein of SARS‑CoV‑2 Induces Endothelial Inflammation through Integrin α5β1 and NF-κB Signaling,” J. Biol. Chem. 2022, 298, 3: 101695. doi: https://doi.org/10.1016/j.jbc.2022.101695
Sharma VK et al., “Nanocurcumin Potently Inhibits SARS‑CoV‑2 Spike Protein‑Induced Cytokine Storm by Deactivation of MAPK/NF-κB Signaling in Epithelial Cells,” ACS Appl. Bio Mater. 2022, 5, 2: 483–491. doi: https://doi.org/10.1021/acsabm.1c00874
Bhattacharyya S and JK Tobacman, “SARS‑CoV‑2 spike protein‑ACE2 interaction increases carbohydrate sulfotransferases and reduces N‑acetylgalactosamine‑4-sulfatase by p38 MAPK,” Signal Transduct Target Ther 2024, 9, 39. doi: https://doi.org/10.1038/s41392‑024‑01741‑3
Q. Mastocytes
Cao JB et al., “Mast cell degranulation‑triggered by SARS‑CoV‑2 induces tracheal‑bronchial epithelial inflammation and injury,” Virol. Sin. 2024, 39, 2: 309‑318. doi: https://doi.org/10.1016/j.virs.2024.03.001
Fajloun Z et al., “SARS‑CoV‑2 or Vaccinal Spike Protein can Induce Mast Cell Activation Syndrome (MCAS),” Infect Disord Drug Targets, 2025, 25, 1: e300424229561. doi: https://doi.org/10.2174/0118715265319896240427045026
Wu ML et al., “Mast cell activation triggered by SARS‑CoV‑2 causes inflammation in brain microvascular endothelial cells and microglia,” Front. Cell. Infect. Microbiol., 2024, 14. doi: https://doi.org/10.3389/fcimb.2024.1358873
R. Microglie
Chang MH et al., “SARS‑CoV‑2 Spike Protein 1 Causes Aggregation of α-Synuclein via Microglia- Induced Inflammation and Production of Mitochondrial ROS: Potential Therapeutic Applications of Metformin,” Biomedicines 2024, 12, 6: 1223. doi: https://doi.org/10.3390/biomedicines12061223
Clough E et al., “Mitochondrial Dynamics in SARS‑COV2 Spike Protein Treated Human Microglia: Implications for Neuro‑COVID,” J. Neuroimmune Pharmacol. 2021, 16, 4: 770–784. doi: https://doi.org/10.1007/s11481‑021‑10015‑6
Frank MG et al., “SARS‑CoV‑2 Spike S1 Subunit Induces Neuroinflammatory, Microglial and Behavioral Sickness Responses: Evidence of PAMP‑Like Properties,” Brain Behav. Immun. 2022, 100: 267277. doi: https://doi.org/10.1016/j.bbi.2021.12.007
Mishra R and AC Banerjea, “SARS‑CoV‑2 Spike targets USP33‑IRF9 axis via exosomal miR‑148a to activate human microglia,” Front. Immunol. 2021, 12: 656700. doi: https://doi.org/10.3389/fimmu.2021.656700
Olajide OA et al., “SARS‑CoV‑2 spike glycoprotein S1 induces neuroinflammation in BV‑2 microglia,” Mol. Neurobiol. 2022, 59: 445‑458. doi: https://doi.org/10.1007/s12035‑021‑02593‑6
Wu ML et al., “Mast cell activation triggered by SARS‑CoV‑2 causes inflammation in brain microvascular endothelial cells and microglia,” Front. Cell. Infect. Microbiol. 2024, 14. doi: https://doi.org/10.3389/fcimb.2024.1358873
S. Microvasculaire
Avolio E et al., “The SARS‑CoV‑2 Spike Protein Disrupts Human Cardiac Pericytes Function through CD147 Receptor‑Mediated Signalling: A Potential Non‑infective Mechanism of COVID‑19 Microvascular Disease,” Clin. Sci. 2021, 135, 24: 2667–2689. doi: https://doi.org/10.1042/CS20210735
Bhargavan B and GD Kanmogne, “SARS‑CoV‑2 spike proteins and cell–cell communication inhibits TFPI and induces thrombogenic factors in human lung microvascular endothelial cells and neutrophils: implications for COVID‑19 coagulopathy pathogenesis,” Int. J. Mol. Sci. 2022, 23, 18: 10436. doi: https://doi.org/10.3390/ijms231810436
Kulkoviene G et al., “Differential Mitochondrial, Oxidative Stress and Inflammatory Responses to SARS‑CoV‑2 Spike Protein Receptor Binding Domain in Human Lung Microvascular, Coronary Artery Endothelial and Bronchial Epithelial Cells,” Int. J. Mol. Sci. 2024, 25, 6: 3188.doi: https://doi.org/10.3390/ijms25063188
Magro N et al., “Disruption of the blood‑brain barrier is correlated with spike endocytosis by ACE2 + endothelia in the CNS microvasculature in fatal COVID‑19. Scientific commentary on ‘Detection of blood‑brain barrier disruption in brains of patients with COVID‑19, but no evidence of brain penetration by SARS‑CoV‑2’,” Acta Neuropathol. 2024, 147, 1: 47. doi: https://doi.org/10.1007/s00401‑023‑02681‑y
Panigrahi S et al., “SARS‑CoV‑2 Spike Protein Destabilizes Microvascular Homeostasis,” Microbiol Spectr. 2021, 9, 3: e0073521. doi: https://doi.org/10.1128/Spectrum.00735‑21
Perico L et al., “SARS‑CoV‑2 Spike Protein 1 Activates Microvascular Endothelial Cells and Complement System Leading to Platelet Aggregation,” Front. Immunol. 2022, 13, 827146. doi: https://doi.org/10.3389/fimmu.2022.827146
Wu ML et al., “Mast cell activation triggered by SARS‑CoV‑2 causes inflammation in brain microvascular endothelial cells and microglia,” Front. Cell. Infect. Microbiol. 2024, 14. doi: https://doi.org/10.3389/fcimb.2024.1358873
Zekri‑Nechar K et al., “Spike Protein Subunits of SARS‑CoV‑2 Alter Mitochondrial Metabolism in Human Pulmonary Microvascular Endothelial Cells: Involvement of Factor Xa,” Dis. Markers 2022: 1118195. doi: https://doi.org/10.1155/2022/1118195
T. Mitochondries/métabolisme
Cao X et al., “The SARS‑CoV‑2 spike protein induces long‑term transcriptional perturbations of mitochondrial metabolic genes, causes cardiac fibrosis, and reduces myocardial contractile in obese mice,” Mol. Metab. 2023, 74, 101756. doi: https://doi.org/10.1016/j.molmet.2023.101756
Chang MH et al., “SARS‑CoV‑2 Spike Protein 1 Causes Aggregation of α-Synuclein via Microglia- Induced Inflammation and Production of Mitochondrial ROS: Potential Therapeutic Applications of Metformin,” Biomedicines 2024, 12, 6: 1223. doi: https://doi.org/10.3390/biomedicines12061223
Clough E et al., “Mitochondrial Dynamics in SARS‑COV2 Spike Protein Treated Human Microglia: Implications for Neuro‑COVID,” Journal of Neuroimmune Pharmacology 2021, 16, 4: 770–784. doi: https://doi.org/10.1007/s11481‑021‑10015‑6
Huynh TV et al., “Spike Protein Impairs Mitochondrial Function in Human Cardiomyocytes: Mechanisms Underlying Cardiac Injury in COVID‑19,” Cells 2023, 12, 877. doi: https://doi.org/10.3390/cells12060877
Kulkoviene G et al., “Differential Mitochondrial, Oxidative Stress and Inflammatory Responses to SARS‑CoV‑2 Spike Protein Receptor Binding Domain in Human Lung Microvascular, Coronary Artery Endothelial and Bronchial Epithelial Cells,” Int. J. Mol. Sci. 2024, 25, 6: 3188.doi: https://doi.org/10.3390/ijms25063188
Mercado‑Gómez M et al., “The spike of SARS‑CoV‑2 promotes metabolic rewiring in hepatocytes,” Commun. Biol. 2022, 5, 827. doi: https://doi.org/10.1038/s42003‑022‑03789‑9
Nguyen V, “The Spike Protein of SARS‑CoV‑2 Impairs Lipid Metabolism and Increases Susceptibility to Lipotoxicity: Implication for a Role of Nrf2,” Cells 2022, 11, 12: 1916. doi: https://doi.org/10.3390/cells11121916
Zekri‑Nechar K et al., “Spike Protein Subunits of SARS‑CoV‑2 Alter Mitochondrial Metabolism in Human Pulmonary Microvascular Endothelial Cells: Involvement of Factor Xa,” Dis. Markers 2022, 1118195. doi: https://doi.org/10.1155/2022/111819
U. Myocardite/cardiomyopathie
Avolio E et al., “The SARS‑CoV‑2 Spike Protein Disrupts Human Cardiac Pericytes Function through CD147 Receptor‑Mediated Signalling: A Potential Non‑infective Mechanism of COVID‑19 Microvascular Disease,” Clin. Sci. 2021, 135, 24: 2667–2689. doi: https://doi.org/10.1042/CS20210735
Baumeier C et al., “Intramyocardial Inflammation after COVID‑19 Vaccination: An Endomyocardial Biopsy‑Proven Case Series,” Int. J. Mol. Sci. 2022, 23: 6940. doi: https://doi.org/10.3390/ijms23136940
Bellavite P et al., “Immune response and molecular mechanisms of cardiovascular adverse effects of spike proteins from SARS‑coV‑2 and mRNA vaccines,” Biomedicines 2023, 11, 2: 451. doi: https://doi.org/10.3390/biomedicines11020451
Boretti A. “PǪǪ Supplementation and SARS‑CoV‑2 Spike Protein‑Induced Heart Inflammation,” Nat. Prod. Commun. 2022, 17, 1934578×221080929. doi: https://doi.org/10.1177/1934578X221080929
Cao X et al., “The SARS‑CoV‑2 spike protein induces long‑term transcriptional perturbations of mitochondrial metabolic genes, causes cardiac fibrosis, and reduces myocardial contractile in obese mice,” Mol. Metab. 2023, 74, 101756. doi: https://doi.org/10.1016/j.molmet.2023.101756
Clemens DJ et al., “SARS‑CoV‑2 spike protein‑mediated cardiomyocyte fusion may contribute to increased arrhythmic risk in COVID‑19,” PLoS One 2023, 18, 3: e0282151.doi: https://doi.org/10.1371/journal.pone.0282151
De Sousa PMB et al., “Fatal Myocarditis following COVID‑19 mRNA Immunization: A Case Report and Differential Diagnosis Review,” Vaccines 2024, 12, 2: 194.doi: https://doi.org/10.3390/vaccines12020194
Forte E, “Circulating spike protein may contribute to myocarditis after COVID‑19 vaccination,” Nat. Cardiovasc. Res. 2023, 2: 100. doi: https://doi.org/10.1038/s44161‑023‑00222‑0
Huang X et al., “Sars‑Cov‑2 Spike Protein‑Induced Damage of hiPSC‑Derived Cardiomyocytes,” Adv. Biol. 2022, 6, 7: e2101327. doi: https://doi.org/10.1002/adbi.202101327
Hulscher N et al., “Autopsy findings in cases of fatal COVID‑19 vaccine‑induced myocarditis,” ESC Heart Failure 2024. doi: https://doi.org/10.1002/ehf2.14680
Huynh TV et al., “Spike Protein Impairs Mitochondrial Function in Human Cardiomyocytes: Mechanisms Underlying Cardiac Injury in COVID‑19,” Cells 2023, 12, 877. doi: https://doi.org/10.3390/cells12060877
Huynh TV et al., “Spike Protein of SARS‑CoV‑2 Activates Cardiac Fibrogenesis through NLRP3 Inflammasomes and NF-κB Signaling,” Cells 2024, 13, 16: 1331.doi: https://doi.org/10.3390/cells13161331
Kato Y et al., “TRPC3‑Nox2 Protein Complex Formation Increases the Risk of SARS‑CoV‑2 Spike Protein‑Induced Cardiomyocyte Dysfunction through ACE2 Upregulation,” Int. J. Mol. Sci. 2023, 24, 1:102. doi: https://doi.org/10.3390/ijms24010102
Lin Z, “More than a key—the pathological roles of SARS‑CoV‑2 spike protein in COVID‑19 related cardiac injury,” Sports Med Health Sci 2023, 6, 3: 209‑220. doi: https://doi.org/10.1016/j.smhs.2023.03.004
Albornoz EA et al., “SARS‑CoV‑2 drives NLRP3 inflammasome activation in human microglia through spike protein,” Mol. Psychiatr. 2023, 28: 2878–2893. doi: https://doi.org/10.1038/s41380‑022‑01831‑0
Arjsri P et al., “Hesperetin from root extract of Clerodendrum petasites S. Moore inhibits SARS‑CoV‑2 spike protein S1 subunit‑induced Nlrp3 inflammasome in A549 lung cells via modulation of the Akt/Mapk/Ap‑1 pathway,” Int. J. Mol. Sci. 2022, 23, 18: 10346. doi: https://doi.org/10.3390/ijms231810346
Chittasupho C et al., “Inhibition of SARS‑CoV‑2-Induced NLRP3 Inflammasome‑Mediated Lung Cell Inflammation by Triphala‑Loaded Nanoparticle Targeting Spike Glycoprotein S1,” Pharmaceutics 2024, 16, 6: 751. https://doi.org/10.3390/pharmaceutics16060751
Chittasupho C et al., “Targeting spike glycoprotein S1 mediated by NLRP3 inflammasome machinery and the cytokine releases in A549 lung epithelial cells by nanocurcumin,” Pharmaceuticals (Basel) 2023, 16, 6: 862. doi: https://doi.org/10.3390/ph16060862
Corpetti C et al., “Cannabidiol inhibits SARS‑Cov‑2 spike (S) protein‑induced cytotoxicity and inflammation through a PPARγ-dependent TLR4/NLRP3/Caspase‑1 signaling suppression in Caco‑2 cell line,” Phytother. Res. 2021, 35, 12: 6893–6903. doi: https://doi.org/10.1002/ptr.7302
Del Re A et al., “Ultramicronized Palmitoylethanolamide Inhibits NLRP3 Inflammasome Expression and Pro‑Inflammatory Response Activated by SARS‑CoV‑2 Spike Protein in Cultured Murine Alveolar Macrophages.” Metabolites 2021, 11, 9: 592. dsoi: https://doi.org/10.3390/metabo11090592
Dissook S et al., “Luteolin‑rich fraction from Perilla frutescens seed meal inhibits spike glycoprotein S1 of SARS‑CoV‑2-induced NLRP3 inflammasome lung cell inflammation via regulation of JAK1/STAT3 pathway: A potential anti‑inflammatory compound against inflammation‑induced long- COVID,” Front. Med. 2023, 9: 1072056. doi: https://doi.org/10.3389/fmed.2022.1072056
Huynh TV et al., “Spike Protein of SARS‑CoV‑2 Activates Cardiac Fibrogenesis through NLRP3 Inflammasomes and NF-κB Signaling,” Cells 2024, 13, 16: 1331.doi: https://doi.org/10.3390/cells13161331
Jiang Ǫ et al., “SARS‑CoV‑2 spike S1 protein induces microglial NLRP3‑dependent neuroinflammation and cognitive impairment in mice,” Exp. Neurol. 2025, 383: 115020. doi: https://doi.org/10.1016/j.expneurol.2024.115020
Kucia M et al. “An evidence that SARS‑Cov‑2/COVID‑19 spike protein (SP) damages hematopoietic stem/progenitor cells in the mechanism of pyroptosis in Nlrp3 inflammasome‑dependent manner,” Leukemia 2021, 35: 3026‑3029. doi: https://doi.org/10.1038/s41375‑021‑01332‑z
Ratajczak MZ et al., “SARS‑CoV‑2 Entry Receptor ACE2 Is Expressed on Very Small CD45- Precursors of Hematopoietic and Endothelial Cells and in Response to Virus Spike Protein Activates the Nlrp3 Inflammasome,” Stem Cell Rev Rep. 2021, 17, 1: 266‑277. doi: https://doi.org/10.1007/s12015‑020‑10010‑z
Semmarath W et al., “Cyanidin‑3-O‑glucoside and Peonidin‑3-O‑glucoside‑Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS‑CoV‑2 Spike Glycoprotein S1- Induction In Vitro in A549 Lung Cells and THP‑1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway,” Nutrients 2022, 14, 13: 2738. doi: https://doi.org/10.3390/nu14132738
Villacampa A et al., “SARS‑CoV‑2 S protein activates NLRP3 inflammasome and deregulates coagulation factors in endothelial and immune cells,” Cell Commun. Signal. 2024, 22, 38. doi: https://doi.org/10.1186/s12964‑023‑01397‑6
W. Oculaire, ophtalmique, conjonctival
Golob‑Schwarzl N et al., “SARS‑CoV‑2 spike protein functionally interacts with primary human conjunctival epithelial cells to induce a pro‑inflammatory response,” Eye 2022, 36: 2353–5. doi: https://doi.org/10.1038/s41433‑022‑02066‑7
Grishma K and Das Sarma, “The Role of Coronavirus Spike Protein in Inducing Optic Neuritis in Mice: Parallels to the SARS‑CoV‑2 Virus,” J Neuroophthalmol 2024, 44, 3: 319‑329. doi: 10.1097/WNO.0000000000002234
Zhu G et al., “SARS‑CoV‑2 spike protein‑induced host inflammatory response signature in human corneal epithelial cells,” Mol. Med. Rep. 2021, 24: 584. doi: https://doi.org/10.3892/mmr.2021.12223
X. Autre signalisation cellulaire
Caohuy H et al., “Inflammation in the COVID‑19 airway is due to inhibition of CFTR signaling by the SARS‑CoV‑2 spike protein,” Sci. Rep. 2024, 14: 16895. doi: https://doi.org/10.1038/s41598‑024‑66473‑4
Choi JY et al., “SARS‑CoV‑2 spike S1 subunit protein‑mediated increase of beta‑secretase 1 (BACE1) impairs human brain vessel cells,” Biochem. Biophys. Res. Commun. 2022, 625, 20: 66‑71.doi: https://doi.org/10.1016/j.bbrc.2022.07.113
Corpetti C et al., “Cannabidiol inhibits SARS‑Cov‑2 spike (S) protein‑induced cytotoxicity and inflammation through a PPARγ-dependent TLR4/NLRP3/Caspase‑1 signaling suppression in Caco‑2 cell line,” Phytother. Res. 2021, 35, 12: 6893–6903. doi: https://doi.org/10.1002/ptr.7302
Gracie NP et al., “Cellular signalling by SARS‑CoV‑2 spike protein,” Microbiology Australia 2024, 45, 1: 13‑17. doi: https://doi.org/10.1071/MA24005
Li F et al., “SARS‑CoV‑2 Spike Promotes Inflammation and Apoptosis Through Autophagy by ROS- Suppressed PI3K/AKT/mTOR Signaling,” Biochim Biophys Acta BBA – Mol Basis Dis 2021, 1867: 166260. doi: https://doi.org/10.1016/j.bbadis.2021.166260
Li K et al., “SARS‑CoV‑2 Spike protein promotes vWF secretion and thrombosis via endothelial cytoskeleton‑associated protein 4 (CKAP4),” Signal Transduct Targ Ther 2022, 7, 332. doi: https://doi.org/10.1038/s41392‑022‑01183‑9
Moutal A et al., “SARS‑CoV‑2 Spike protein co‑opts VEGF‑A/Neuropilin‑1 receptor signaling to induce analgesia,” Pain 2020, 162, 1: 243–252. doi: https://doi.org/10.1097/j.pain.0000000000002097
Munavilli GG et al., “COVID‑19/SARS‑CoV‑2 virus spike protein‑related delayed inflammatory reaction to hyaluronic acid dermal fillers: a challenging clinical conundrum in diagnosis and treatment,” Arch. Dermatol. Res. 2022, 314: 1‑15. doi: https://doi.org/10.1007/s00403‑021‑02190‑6
Prieto‑Villalobos J et al., “SARS‑CoV‑2 spike protein S1 activates Cx43 hemichannels and disturbs intracellular Ca2+ dynamics,” Biol Res. 2023, 56, 1: 56. doi: https://doi.org/10.1186/s40659‑023‑00468‑9
Rotoli BM et al., “Endothelial cell activation by SARS‑CoV‑2 spike S1 protein: A crosstalk between endothelium and innate immune cells,” Biomedicines 2021, 9, 9: 1220. doi: https://doi.org/10.3390/biomedicines9091220
Singh N and Anuradha Bharara Singh, “S2 Subunit of SARS‑nCoV‑2 Interacts with Tumor Suppressor Protein p53 and BRCA: An in Silico Study,” Translational Oncology 2020, 13, 10: 100814. doi: https://doi.org/10.1016/j.tranon.2020.100814
Singh RD, “The spike protein of sars‑cov‑2 induces heme oxygenase‑1: pathophysiologic implications,” Biochim Biophys Acta, Mol Basis Dis 2022, 1868, 3: 166322. doi: https://doi.org/10.1016/j.bbadis.2021.166322
Solis O et al., “The SARS‑CoV‑2 spike protein binds and modulates estrogen receptors,” Sci. Adv. 2022, 8, 48: eadd4150. doi: https://doi.org/10.1126/sciadv.add4150
Suzuki YJ et al., “SARS‑CoV‑2 spike protein‑mediated cell signaling in lung vascular cells,” Vascul. Pharmacol. 2021, 137: 106823. doi: https://doi.org/10.1016/j.vph.2020.106823
Suzuki YJ and SG Gychka, “SARS‑CoV‑2 Spike Protein Elicits Cell Signaling in Human Host Cells: Implications for Possible Consequences of COVID‑19 Vaccines,” Vaccines 2021, 9, 1: 36. doi: https://doi.org/10.3390/vaccines9010036
Tillman TS et al., “SARS‑CoV‑2 Spike Protein Downregulates Cell Surface alpha7nAChR through a Helical Motif in the Spike Neck,” ACS Chem. Neurosci. 2023, 14, 4: 689–698. doi: https://doi.org/10.1021/acschemneuro.2c00610
Y. Grossesse
Erdogan MA, “Prenatal SARS‑CoV‑2 Spike Protein Exposure Induces Autism‑Like Neurobehavioral Changes in Male Neonatal Rats,” J Neuroimmune Pharmacol. 2023, 18, 4: 573‑591. doi: https://doi.org/10.1007/s11481‑023‑10089‑4
Guo X et al., “Regulation of proinflammatory molecules and tissue factor by SARS‑CoV‑2 spike protein in human placental cells: implications for SARS‑CoV‑2 pathogenesis in pregnant women,” Front. Immunol. 2022, 13: 876555–876555. doi: https://doi.org/10.3389/fimmu.2022.876555
Kammala AK et al., “In vitro mRNA‑S maternal vaccination induced altered immune regulation at the maternal‑fetal interface,” Am. J. Reprod. Immunol. 2024, 91, 5: e13861. doi: https://doi.org/10.1111/aji.13861
Z. Pulmonaire, respiratoire
Bhargavan B and GD Kanmogne, “SARS‑CoV‑2 spike proteins and cell–cell communication inhibits TFPI and induces thrombogenic factors in human lung microvascular endothelial cells and neutrophils: implications for COVID‑19 coagulopathy pathogenesis,” Int. J. Mol. Sci. 2022, 23, 18: 10436. doi: https://doi.org/10.3390/ijms231810436
Biancatelli RMLC et al., “The SARS‑CoV‑2 spike protein subunit S1 induces COVID‑19‑like acute lung injury in Kappa18‑hACE2 transgenic mice and barrier dysfunction in human endothelial cells,” Am. J.Physiol. Lung Cell. Mol. Physiol. 2021, 321: L477–L484. doi: https://doi.org/10.1152/ajplung.00223.2021
Cao JB et al., “Mast cell degranulation‑triggered by SARS‑CoV‑2 induces tracheal‑bronchial epithelial inflammation and injury,” Virol. Sin. 2024, 39, 2: 309‑318. doi: https://doi.org/10.1016/j.virs.2024.03.001
Cao X et al., “Spike protein of SARS‑CoV‑2 activates macrophages and contributes to induction of acute lung inflammation in male mice,” FASEB J. 2021, 35, e21801. doi: https://doi.org/10.1096/fj.202002742RR
Caohuy H et al., “Inflammation in the COVID‑19 airway is due to inhibition of CFTR signaling by the SARS‑CoV‑2 spike protein,” Sci. Rep. 2024, 14: 16895. doi: https://doi.org/10.1038/s41598‑024‑66473‑4
Chittasupho C et al., “Inhibition of SARS‑CoV‑2-Induced NLRP3 Inflammasome‑Mediated Lung Cell Inflammation by Triphala‑Loaded Nanoparticle Targeting Spike Glycoprotein S1,” Pharmaceutics 2024, 16, 6: 751. https://doi.org/10.3390/pharmaceutics16060751
Chittasupho C et al., “Targeting spike glycoprotein S1 mediated by NLRP3 inflammasome machinery and the cytokine releases in A549 lung epithelial cells by nanocurcumin,” Pharmaceuticals (Basel) 2023, 16, 6: 862. doi: https://doi.org/10.3390/ph16060862
Del Re A et al., “Intranasal delivery of PEA‑producing Lactobacillus paracasei F1S alleviates SARS- CoV‑2 spike protein‑induced lung injury in mice,” Transl. Med. Commun. 2024, 9, 9. doi: https://doi.org/10.1186/s41231‑024‑00167‑x
Forsyth CB et al., “The SARS‑CoV‑2 S1 spike protein promotes MAPK and NF‑kB activation in human lung cells and inflammatory cytokine production in human lung and intestinal epithelial cells,” Microorganisms 2022, 10, 10: 1996. doi: https://doi.org/10.3390/microorganisms10101996
Greenberger JS et al., “SARS‑CoV‑2 Spike Protein Induces Oxidative Stress and Senescence in Mouse and Human Lung,” In Vivo 2024, 38, 4: 1546‑1556; doi: https://doi.org/10.21873/invivo.13605
Jana S et al., “Cell‑free hemoglobin does not attenuate the effects of SARS‑CoV‑2 spike protein S1 subunit in pulmonary endothelial cells,” Int. J. Mol. Sci. 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Kulkoviene G et al., “Differential Mitochondrial, Oxidative Stress and Inflammatory Responses to SARS‑CoV‑2 Spike Protein Receptor Binding Domain in Human Lung Microvascular, Coronary Artery Endothelial and Bronchial Epithelial Cells,” Int. J. Mol. Sci. 2024, 25, 6: 3188. doi: https://doi.org/10.3390/ijms25063188
Liang S et al., “SARS‑CoV‑2 spike protein induces IL‑18‑mediated cardiopulmonary inflammation via reduced mitophagy,” Signal Transduct Target Ther 2023, 8, 103. doi: https://doi.org/10.1038/s41392‑023‑01368‑w
Liu T et al., “RS‑5645 attenuates inflammatory cytokine storm induced by SARS‑CoV‑2 spike protein and LPS by modulating pulmonary microbiota,” Int J Biol Sci. 2021, 17, 13: 3305–3319. doi: https://doi.org/10.7150/ijbs.63329
Palestra F et al. “SARS‑CoV‑2 Spike Protein Activates Human Lung Macrophages,” Int. J. Mol. Sci. 2023, 24, 3: 3036. doi: https://doi.org/10.3390/ijms24033036
Park C et al., “Murine alveolar Macrophages Rapidly Accumulate intranasally Administered SARS- CoV‑2 Spike Protein leading to neutrophil Recruitment and Damage,” Elife 2024, 12: RP86764. doi: https://doi.org/10.7554/eLife.86764.3
Puthia MTL et al., “Experimental model of pulmonary inflammation induced by SARS‑CoV‑2 spike protein and endotoxin,” ACS Pharmacol Transl Sci. 2022, 5, 3: 141–8. doi: https://doi.org/10.1021/acsptsci.1c00219
Rahman M et al., “Differential Effect of SARS‑CoV‑2 Spike Glycoprotein 1 on Human Bronchial and Alveolar Lung Mucosa Models: Implications for Pathogenicity,” Viruses 2021, 13, 12: 2537. doi: https://doi.org/10.3390/v13122537
Ruben ML et al., “The SARS‑CoV‑2 spike protein subunit S1 induces COVID‑19‑like acute lung injury in Κ18‑hACE2 transgenic mice and barrier dysfunction in human endothelial cells,” Am J Physiol Lung Cell Mol Physiol. 2021, 321, 2: L477‑L484. doi: https://doi.org/10.1152/ajplung.00223.2021
Segura‑Villalobos D et al., “Jacareubin inhibits TLR4‑induced lung inflammatory response caused by the RBD domain of SARS‑CoV‑2 Spike protein,” Pharmacol. Rep. 2022, 74: 1315–1325. doi: https://doi.org/10.1007/s43440‑022‑00398‑5
Semmarath W et al., “Cyanidin‑3-O‑glucoside and Peonidin‑3-O‑glucoside‑Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS‑CoV‑2 Spike Glycoprotein S1- Induction In Vitro in A549 Lung Cells and THP‑1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway,” Nutrients 2022, 14, 13: 2738. doi: https://doi.org/10.3390/nu14132738
Sirsendu J et al., “Cell‑Free Hemoglobin Does Not Attenuate the Effects of SARS‑CoV‑2 Spike Protein S1 Subunit in Pulmonary Endothelial Cells,” Int. J. Mol. Sci. 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Sui Y et al., “SARS‑CoV‑2 Spike Protein Suppresses ACE2 and Type I Interferon Expression in Primary Cells From Macaque Lung Bronchoalveolar Lavage,” Front. Immunol. 2021, 12. doi: https://doi.org/10.3389/fimmu.2021.658428
Sung PS et al., “CLEC5A and TLR2 Are Critical in SARS‑CoV‑2-Induced NET Formation and Lung Inflammation,” J. Biomed. Sci. 2022, 29, 52. doi: https://doi.org/10.1186/s12929‑022‑00832‑z
Suzuki YJ et al., “SARS‑CoV‑2 spike protein‑mediated cell signaling in lung vascular cells,” Vascul. Pharmacol. 2021, 137: 106823. doi: https://doi.org/10.1016/j.vph.2020.106823
Zekri‑Nechar K et al., “Spike Protein Subunits of SARS‑CoV‑2 Alter Mitochondrial Metabolism in Human Pulmonary Microvascular Endothelial Cells: Involvement of Factor Xa,” Dis. Markers 2022, 1118195. doi: https://doi.org/10.1155/2022/1118195
AA. Système rénine‑angiotensine‑aldostérone
Lehmann KJ, “SARS‑CoV‑2-Spike Interactions with the Renin‑Angiotensin‑Aldosterone System – Consequences of Adverse Reactions of Vaccination,” J Biol Today’s World 2023, 12/4: 001‑013. doi: https://doi.org/10.31219/osf.io/27g5h
Matsuzawa Y et al., “Impact of Renin–Angiotensin–Aldosterone System Inhibitors on COVID‑19,” Hypertens. Res. 2022, 45, 7: 1147–1153, doi: https://doi.org/10.1038/s41440‑022‑00922‑3
AB. Sénescence/vieillissement
Duarte C, “Age‑dependent effects of the recombinant spike protein/SARS‑CoV‑2 on the M‑CSF- and IL‑34‑differentiated macrophages in vitro,” Biochem. Biophys. Res. Commun. 2021, 546: 97–102. doi: https://doi.org/10.1016/j.bbrc.2021.01.104
Greenberger JS et al., “SARS‑CoV‑2 Spike Protein Induces Oxidative Stress and Senescence in Mouse and Human Lung,” In Vivo 2024, 38, 4: 1546‑1556. doi: https://doi.org/10.21873/invivo.13605
Meyer K et al., “SARS‑CoV‑2 Spike Protein Induces Paracrine Senescence and Leukocyte Adhesion in Endothelial Cells,” J. Virol. 2021, 95, e0079421. doi: https://doi.org/10.1128/jvi.00794‑21
AC. Cellules souches
Balzanelli MG et al., “The Role of SARS‑CoV‑2 Spike Protein in Long‑term Damage of Tissues and Organs, the Underestimated Role of Retrotransposons and Stem Cells, a Working Hypothesis,” Endocr Metab Immune Disord Drug Targets 2025, 25, 2: 85‑98. doi: https://doi.org/10.2174/0118715303283480240227113401
Kucia M et al. “An evidence that SARS‑Cov‑2/COVID‑19 spike protein (SP) damages hematopoietic stem/progenitor cells in the mechanism of pyroptosis in Nlrp3 inflammasome‑dependent manner,” Leukemia 2021, 35: 3026‑3029. doi: https://doi.org/10.1038/s41375‑021‑01332‑z
Ropa J et al., “Human Hematopoietic Stem, Progenitor, and Immune Cells Respond Ex Vivo to SARS- CoV‑2 Spike Protein,” Stem Cell Rev Rep. 2021, 17, 1: 253‑265. doi: https://doi.org/10.1007/s12015‑020‑10056‑z
Cattin-Ortolá J et al., “Sequences in the cytoplasmic tail of SARS-CoV-2 Spike facilitate expression at the cell surface and syncytia formation,” Nat Commun 2021, 12, 1: 5333. doi: https://doi.org/10.1038/s41467‑021‑25589‑1
Clemens DJ et al., “SARS‑CoV‑2 spike protein‑mediated cardiomyocyte fusion may contribute to increased arrhythmic risk in COVID‑19,” PLoS One 2023, 18, 3: e0282151. doi: https://doi.org/10.1371/journal.pone.0282151
Lazebnik Y, “Cell fusion as a link between the SARS‑CoV‑2 spike protein, COVID‑19 complications, and vaccine side effects,” Oncotarget 2021, 12, 25: 2476‑2488. doi: https://doi.org/10.18632/oncotarget.28088
Liu X et al., “SARS‑CoV‑2 spike protein‑induced cell fusion activates the cGAS‑STING pathway and the interferon response,” Sci Signal. 2022, 15, 729: eabg8744. doi: https://doi.org/10.1126/scisignal.abg8744
Martinez‑Marmol R et al., “SARS‑CoV‑2 infection and viral fusogens cause neuronal and glial fusion that compromises neuronal activity,” Sci. Adv. 2023, 9, 23. doi: https://doi.org/10.1126/sciadv.adg2248
Rajah MM et al., “SARS‑CoV‑2 Alpha, Beta, and Delta variants display enhanced spike‑mediated syncytia formation,” EMBO J. 2021, 40: e108944. doi: https://doi.org/10.15252/embj.2021108944
Shirato K and Takako Kizaki, “SARS‑CoV‑2 Spike Protein S1 Subunit Induces Pro- inflammatory Responses via Toll‑Like Receptor 4 Signaling in Murine and Human Macrophages,” Heliyon 2021, 7, 2: e06187. doi: https://doi.org/10.1016/j.heliyon.2021.e06187
Theuerkauf SA et al., “Ǫuantitative assays reveal cell fusion at minimal levels of SARS‑CoV‑2 spike protein and fusion from without,” iScience 2021, 24, 3: 102170. doi: https://doi.org/10.1016/j.isci.2021.102170
Zhang Z et al., “SARS‑CoV‑2 spike protein dictates syncytium‑mediated lymphocyte elimination,” Cell Death Differ. 2021, 28: 2765–2777. doi: https://doi.org/10.1038/s41418‑021‑00782‑3
AE. Thérapeutiques
Almehdi AM et al., “SARS‑CoV‑2 Spike Protein: Pathogenesis, Vaccines, and Potential Therapies,” Infection 2021, 49, 5: 855–876. doi: https://doi.org/10.1007/s15010‑021‑01677‑8
Boretti A, “PǪǪ Supplementation and SARS‑CoV‑2 Spike Protein‑Induced Heart Inflammation,” Nat. Prod. Commun. 2022, 17, 1934578×221080929. doi: https://doi.org/10.1177/1934578X221080929
Boschi C et al., “SARS‑CoV‑2 Spike Protein Induces Hemagglutination: Implications for COVID‑19 Morbidities and Therapeutics and for Vaccine Adverse Effects,” Int. J. Biol. Macromol. 2022, 23, 24: 15480. doi: https://doi.org/10.3390/ijms232415480
Chang MH et al., “SARS‑CoV‑2 Spike Protein 1 Causes Aggregation of α-Synuclein via Microglia- Induced Inflammation and Production of Mitochondrial ROS: Potential Therapeutic Applications of Metformin,” Biomedicines 2024, 12, 6: 1223. doi: https://doi.org/10.3390/biomedicines12061223
Chittasupho C et al., “Inhibition of SARS‑CoV‑2-Induced NLRP3 Inflammasome‑Mediated Lung Cell Inflammation by Triphala‑Loaded Nanoparticle Targeting Spike Glycoprotein S1,” Pharmaceutics 2024, 16, 6: 751. doi: https://doi.org/10.3390/pharmaceutics16060751
Chittasupho C et al., “Targeting spike glycoprotein S1 mediated by NLRP3 inflammasome machinery and the cytokine releases in A549 lung epithelial cells by nanocurcumin,” Pharmaceuticals (Basel) 2023, 16, 6: 862. doi: https://doi.org/10.3390/ph16060862
Corpetti C et al., “Cannabidiol inhibits SARS‑Cov‑2 spike (S) protein‑induced cytotoxicity and inflammation through a PPARγ-dependent TLR4/NLRP3/Caspase‑1 signaling suppression in Caco‑2 cell line,” Phytother. Res. 2021, 35, 12: 6893–6903. doi: https://doi.org/10.1002/ptr.7302
Cory TJ et al., “Metformin Suppresses Monocyte Immunometabolic Activation by SARS‑CoV‑2 Spike Protein Subunit 1,” Front. Immunol. 2021, 12 (Sec. Cytokines and Soluble Mediators in Immunity): 733921. doi: https://doi.org/10.3389/fimmu.2021.733921
Del Re A et al., “Intranasal delivery of PEA‑producing Lactobacillus paracasei F1S alleviates SARS- CoV‑2 spike protein‑induced lung injury in mice,” Transl. Med. Commun. 2024, 9, 9. doi: https://doi.org/10.1186/s41231‑024‑00167‑x
Del Re A et al., “Ultramicronized Palmitoylethanolamide Inhibits NLRP3 Inflammasome Expression and Pro‑Inflammatory Response Activated by SARS‑CoV‑2 Spike Protein in Cultured Murine Alveolar Macrophages,” Metabolites 2021, 11, 9: 592. doi: https://doi.org/10.3390/metabo11090592
Ferrer MD et al., “Nitrite Attenuates the In Vitro Inflammatory Response of Immune Cells to the SARS‑CoV‑2 S Protein without Interfering in the Antioxidant Enzyme Activation,” Int. J. Mol.Sci. 2024, 25, 5: 3001. https://doi.org/10.3390/ijms25053001
Frank MG et al., “SARS‑CoV‑2 S1 subunit produces a protracted priming of the neuroinflammatory, physiological, and behavioral responses to a remote immune challenge: A role for corticosteroids,” Brain Behav. Immun. 2024, 121: 87‑103. doi: https://doi.org/10.1016/j.bbi.2024.07.034
Frühbeck G et al., “FNDC4 and FNDC5 reduce SARS‑CoV‑2 entry points and spike glycoprotein S1- induced pyroptosis, apoptosis, and necroptosis in human adipocytes,” Cell Mol Immunol. 2021, 18, 10: 2457–9. doi: https://doi.org/10.1038/s41423‑021‑00762‑0
Gasparello J et al., “Sulforaphane inhibits the expression of interleukin‑6 and interleukin‑8 induced in bronchial epithelial IB3‑1 cells by exposure to the SARS‑CoV‑2 Spike protein,” Phytomedicine 2021, 87: 153583. doi: https://doi.org/10.1016/j.phymed.2021.153583
Halma MTJ et al., “Exploring autophagy in treating SARS‑CoV‑2 spike protein‑related pathology,” Endocrinol Metab (EnM) 2024, 14: 100163. doi: https://doi.org/10.1016/j.endmts.2024.100163
Jana S et al., “Cell‑free hemoglobin does not attenuate the effects of SARS‑CoV‑2 spike protein S1 subunit in pulmonary endothelial cells,” Int. J. Mol. Sci. 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Jugler C et al., “SARS‑CoV‑2 Spike Protein‑Induced Interleukin 6 Signaling Is Blocked by a Plant- Produced Anti‑Interleukin 6 Receptor Monoclonal Antibody,” Vaccines 2021, 9, 11: 1365. doi: https://doi.org/10.3390/vaccines9111365
Ken W et al., “Low dose radiation therapy attenuates ACE2 depression and inflammatory cytokines induction by COVID‑19 viral spike protein in human bronchial epithelial cells,” Int J Radiat Biol. 2022, 98, 10:1532‑1541. doi: https://doi.org/10.1080/09553002.2022.2055806
Kumar N et al., “SARS‑CoV‑2 spike protein S1‑mediated endothelial injury and pro‑inflammatory state Is amplified by dihydrotestosterone and prevented by mineralocorticoid antagonism,” Viruses 2021, 13, 11: 2209. doi: https://doi.org/10.3390/v13112209
Liu T et al., “RS‑5645 attenuates inflammatory cytokine storm induced by SARS‑CoV‑2 spike protein and LPS by modulating pulmonary microbiota,” Int J Biol Sci. 2021, 17, 13: 3305–3319. doi: https://doi.org/10.7150/ijbs.63329
Loh D, “The potential of melatonin in the prevention and attenuation of oxidative hemolysis and myocardial injury from cd147 SARS‑CoV‑2 spike protein receptor binding,” Melatonin Research 2020, 3, 3: 380‑416. doi: https://doi.org/10.32794/mr11250069
Loh JT et al., “Dok3 restrains neutrophil production of calprotectin during TLR4 sensing of SARS‑CoV- 2 spike protein,” Front. Immunol. 2022, 13 (Sec. Molecular Innate Immunity). doi: https://doi.org/10.3389/fimmu.2022.996637
Marrone L et al., “Tirofiban prevents the effects of SARS‑CoV‑2 spike protein on macrophage activation and endothelial cell death,” Heliyon 2024, 10, 15: e35341. doi: https://doi.org/10.1016/j.heliyon.2024.e35341
Norris B et al., “Evaluation of Glutathione in Spike Protein of SARS‑CoV‑2 Induced Immunothrombosis and Cytokine Dysregulation,” Antioxidants 2024, 13, 3: 271. doi: https://doi.org/10.3390/antiox13030271
Olajide OA et al., “Induction of Exaggerated Cytokine Production in Human Peripheral Blood Mononuclear Cells by a Recombinant SARS‑CoV‑2 Spike Glycoprotein S1 and Its Inhibition by Dexamethasone,” Inffammation 2021, 44: 1865–1877. doi: https://doi.org/10.1007/s10753‑021‑01464‑5
Petrosino S and N Matende, “Elimination/Neutralization of COVID‑19 Vaccine‑Produced Spike Protein: Scoping Review,” Mathews Journal of Nutrition & Dietetics 2024, 7, 2. doi: https://doi.org/10.30654/MJND.10034
Satta S et al., “An engineered nano‑liposome‑human ACE2 decoy neutralizes SARS‑CoV‑2 Spike protein‑induced inflammation in both murine and human macrophages,” Theranostics 2022, 12, 6: 2639–2657. doi: https://doi.org/10.7150/thno.66831
Segura‑Villalobos D et al., “Jacareubin inhibits TLR4‑induced lung inflammatory response caused by the RBD domain of SARS‑CoV‑2 Spike protein,” Pharmacol. Rep. 2022, 74: 1315–1325. doi: https://doi.org/10.1007/s43440‑022‑00398‑5
Semmarath W et al., “Cyanidin‑3-O‑glucoside and Peonidin‑3-O‑glucoside‑Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS‑CoV‑2 Spike Glycoprotein S1- Induction In Vitro in A549 Lung Cells and THP‑1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway,” Nutrients 2022, 14, 13: 2738. doi: https://doi.org/10.3390/nu14132738
Suprewicz L et al., “Recombinant human plasma gelsolin reverses increased permeability of the blood‑brain barrier induced by the spike protein of the SARS‑CoV‑2 virus,” J Neuroinffammation 2022, 19, 1: 282. doi: https://doi.org/10.1186/s12974‑022‑02642‑4
Vargas‑Castro R et al., “Calcitriol prevents SARS‑CoV spike‑induced inflammation in human trophoblasts through downregulating ACE2 and TMPRSS2 expression,” J Steroid Biochem Mol Biol 2025, 245: 106625. doi: https://doi.org/10.1016/j.jsbmb.2024.106625
Youn JY et al., “Therapeutic application of estrogen for COVID‑19: Attenuation of SARS‑CoV‑2 spike protein and IL‑6 stimulated, ACE2‑dependent NOX2 activation, ROS production and MCP‑1 upregulation in endothelial cells,” Redox Biol. 2021, 46: 102099. doi: https://doi.org/10.1016/j.redox.2021.102099
Yu J et al., “Direct activation of the alternative complement pathway by SARS‑CoV‑2 spike proteins is blocked by factor D inhibition,” Blood 2020, 136, 18: 2080–2089. doi: https://doi.org/10.1182/blood.2020008248
AF. Récepteurs Toll‑like (TLRs)
Aboudounya MM and RJ Heads, “COVID‑19 and Toll‑Like Receptor 4 (TLR4): SARS‑CoV‑2 May Bind and Activate TLR4 to Increase ACE2 Expression, Facilitating Entry and Causing Hyperinflammation,” Mediators Inffamm. 2021: 8874339. doi: https://doi.org/10.1155/2021/8874339
Burnett FN et al., “SARS‑CoV‑2 Spike Protein Intensifies Cerebrovascular Complications in Diabetic hACE2 Mice through RAAS and TLR Signaling Activation,” Int. J. Mol. Sci. 2023, 24, 22: 16394. doi: https://doi.org/10.3390/ijms242216394
Carnevale R et al., “Toll‑Like Receptor 4‑Dependent Platelet‑Related Thrombosis in SARS‑CoV‑2 Infection,” Circ. Res. 2023, 132, 3: 290– 305. doi: https://doi.org/10.1161/CIRCRESAHA.122.321541
Corpetti C et al., “Cannabidiol inhibits SARS‑Cov‑2 spike (S) protein‑induced cytotoxicity and inflammation through a PPARγ-dependent TLR4/NLRP3/Caspase‑1 signaling suppression in Caco‑2 cell line,” Phytother. Res. 2021, 35, 12: 6893–6903. doi: https://doi.org/10.1002/ptr.7302
Fontes‑Dantas FL, “SARS‑CoV‑2 Spike Protein Induces TLR4‑Mediated Long- Term Cognitive Dysfunction Recapitulating Post‑COVID‑19 Syndrome in Mice,” Cell Reports 2023, 42, 3: 112189, doi: https://doi.org/10.1016/j.celrep.2023.112189 Khan S et al., “SARS‑CoV‑2 Spike Protein Induces
Kim MJ et al., “The SARS-CoV-2 spike protein induces lung cancer migration and invasion in a TLR2- dependent manner,” Cancer Commun (London) 2023, 44, 2: 273–277. doi: https://doi.org/10.1002/cac2.12485
Kircheis R and O Planz, “Could a Lower Toll‑like Receptor (TLR) and NF-κB Activation Due to a Changed Charge Distribution in the Spike Protein Be the Reason for the Lower Pathogenicity of Omicron?” Int. J. Mol. Sci. 2022, 23, 11: 5966. doi: https://doi.org/10.3390/ijms23115966
Loh JT et al., “Dok3 restrains neutrophil production of calprotectin during TLR4 sensing of SARS‑CoV- 2 spike protein,” Front. Immunol. 2022, 13 (Sec. Molecular Innate Immunity). doi: https://doi.org/10.3389/fimmu.2022.996637
Segura‑Villalobos D et al., “Jacareubin inhibits TLR4‑induced lung inflammatory response caused by the RBD domain of SARS‑CoV‑2 Spike protein,” Pharmacol. Rep. 2022, 74: 1315–1325. doi: https://doi.org/10.1007/s43440‑022‑00398‑5
Sirsendu J et al., “Cell‑Free Hemoglobin Does Not Attenuate the Effects of SARS‑CoV‑2 Spike Protein S1 Subunit in Pulmonary Endothelial Cells,” Int. J. Mol. Sci., 2021, 22, 16: 9041. doi: https://doi.org/10.3390/ijms22169041
Sung PS et al., “CLEC5A and TLR2 Are Critical in SARS‑CoV‑2-Induced NET Formation and Lung Inflammation,” J. Biomed. Sci. 2022, 29, 52. doi: https://doi.org/10.1186/s12929‑022‑00832‑z
Zaki H and S Khan, “SARS‑CoV‑2 spike protein induces inflammatory molecules through TLR2 in macrophages and monocytes,” J. Immunol. 2021, 206 (1_supplement): 62.07. doi: https://doi.org/10.4049/jimmunol.206.Supp.62.07
LETTRE OUVERTE Au Premier ministre François Bayrou, A la ministre de l’Agriculture Annie Genevard,
Monsieur le Premier ministre, Madame la ministre de l’Agriculture,
L’Union européenne s’apprête à modifier radicalement et sans aucune possibilité de retour en arrière notre agriculture et notre alimentation !
Sous pression des lobbys, la Commission européenne pourrait autoriser d’ici deux mois une nouvelle génération d’OGM à envahir nos champs et nos assiettes sans avoir à se plier aux mesures indispensables de prévention sanitaire et environnementale en vigueur depuis plus de vingt ans.
Cette loi, si elle est adoptée, constituerait un recul historique de nos droits et une mise en danger inédite des écosystèmes :
> il n’y aurait plus AUCUNE évaluation des risques que ces nouveaux produits pourraient faire peser sur les abeilles, l’environnement, et la santé humaine, malgré l’alerte des scientifiques et de l’ANSES, l’agence de sécurité sanitaire française, qui reconnaissent formellement l’existence de dangers potentiels pour l’environnement et notre santé ;
> il n’y aurait plus aucun étiquetage, aucune traçabilité et donc aucun moyen pour nous de savoir si les aliments que nous achetons en magasin contiennent des OGM ;
> il n’y aurait plus aucun rempart contre l’emprise des industriels de l’agrochimie sur le système agricole puisque les multinationales accapareront une part grandissante du vivant à coups de milliers de brevets et contamineront les alternatives agroécologiques, qui sont pourtant notre seul espoir face à la catastrophe écologique en cours.
Pour toutes ces raisons, la France doit s’opposer à ce texte dès maintenant.
Si nous, citoyennes et citoyens français, vous écrivons aujourd’hui, c’est parce que le rôle de la France est déterminant dans le cycle de négociations en cours au Conseil de l’Union européenne. Votre vote pourrait immédiatement mettre fin à cette folie.
C’est la seule voie possible et entendable pour protéger notre agriculture et nos agriculteurs, la biodiversité et les droits fondamentaux des citoyens.
C’est pourquoi nous, citoyens, agriculteurs, distributeurs et consommateurs, vous demandons d’adopter une position ferme lors des discussions à venir, contre toute tentative de soustraire les nouveaux OGM aux réglementations européennes existantes sur les OGM, afin de garantir la sécurité de nos aliments, la préservation de la biodiversité et notre liberté de choix.
Vous priant d’agréer, Monsieur le Premier ministre, Madame la ministre, l’expression de notre plus haute considération.
65 943 ont signé. Passons à 100 000Prénom *Nom de familleCourriel *
2 Département de génétique évolutive et de biosystème, Faculté de biologie, Université de Gdansk, Gdansk, Pologne
3 Norwich Medical School, Université d’East Anglia, Norwich, Royaume-Uni
INTRODUCTION: Les masques ont été largement mandatés lors de la récente pandémie SARS-COV-2. En particulier, l’utilisation par la population générale est associée à un risque plus élevé de manipulation incorrecte du masque et de contamination et de conséquences microbiologiques défavorables potentielles.
Méthodes: Nous avons étudié et quantifié l’accumulation bactérienne dans les masques utilisées par la population générale, en utilisant l’ARNr 16S (séquençage Sanger), la culture et l’analyse biochimique ainsi que la coloration au bengale rose. De plus, un aperçu systématique de la littérature sur la contamination du masque facial a été entreprise.
Résultats: Nous avons trouvé une charge bactérienne moyenne de 4,24 × 10 4 CFU a récupéré / masque, avec une charge maximale de 2,85 × 10 5 CFU. Ce maximum est 310 fois supérieur à la valeur limite de la contamination des surfaces de sortie du système de ventilation spécifiées par l’identification allemande VDI 6022. L’identification biochimique et moléculaire a été principalement trouvée des espèces de Staphylococcus (80%), y compris Staphylococcus aureus , ainsi que Bacillus forming. Les rapports de littérature indiquent également la contamination des masques par des opportunistes bactériens et fongiques des genres Acinetobacter, Aspergillus, Alternaria, Bacillus, Cadosporium, Candida, Escherichia, Enterobacter, Enteroccus, Klebsiella (y compris le K. PseudoMOe , STAPHYLOCCUS, MICROSPORUM, MUCOR, MUCOR, PNEUDOMONAS, STAPHYLOCH et Streptococcus . Les dénombrements bactériens augmentent linéairement avec la durée du port.
Discussion: Une utilisation prolongée peut affecter la peau et les microbiomes respiratoires, favorisant les conditions d’œil, de peau, orale et des voies respiratoires consécutives. Ces aspects soulignent le besoin urgent de recherches supplémentaires et une analyse risque-avantage en ce qui concerne l’utilisation du masque, en particulier compte tenu de leur efficacité non prouvée dans la perturbation de la transmission des virus respiratoires et de leurs conséquences sociales indésirables.
Introduction
Les masques de masque couvrant les entrées des Airways ont été largement mandatés lors de la récente pandémie SARS-COV-2, non seulement pour les travailleurs de la santé mais aussi pour la population générale ( 1 ). Les professions ayant des contacts humains fréquents ont été obligés de les porter pendant de longues périodes, tout comme les écoliers ( 1 à 6 ).
Cela soulève des préoccupations raisonnables: premièrement, car l’utilisation par la population générale est associée à un risque plus élevé de manipulation inappropriée du masque ( 7 – 11 ); Deuxièmement parce que leur efficacité contre les infections virales respiratoires n’est pas prouvée par des essais de haute qualité, qui indiquent peu ou pas d’effet ( 12 , 13 ) et troisièmement, car les masques sont supposés seulement avoir des effets positifs ( 14 – 16 ). En réalité, il existe de solides preuves que les masques posent divers risques, en particulier pour les femmes enceintes, les enfants et les adolescents, ainsi que pour les adultes plus âgés et les malades ( 14 , 16 – 19 ). Ils ont plusieurs effets défavorables manifestement, affectant la physiologie ( 14 , 16 , 19-23 16 et, plus évidemment, les ), la psychologie ( , 24 ) interactions sociales ( 25 – 35 ). Les effets sur le développement de l’enfance sont une préoccupation particulière. Ces effets indésirables ont été récemment résumés comme le syndrome d’épuisement induit par le masque ( 14 , 16 , 19 ). Fait intéressant, Spira ( 36 ) et Fögen ( 37 ) ont trouvé des taux d’infection et de mortalité SARS-COV-2 significativement plus élevés dans les cohortes portant le masque: les explications sont incertaines, mais le piégeage viral et le recyclage sont plausibles.
Une autre préoccupation, englobée au sein des MIES, concerne les conséquences microbiologiques défavorables potentielles du port de masques de visage. En raison de la création d’un micro-environnement chaud et humide ( 38 – 41 ), des bactéries, des champignons et même des virus peuvent s’accumuler des deux côtés des masques usés ( 42 – 46 ). Jusqu’à présent, ces aspects n’ont pas été évalués en profondeur. Le but de notre étude pilote était d’évaluer, de visualiser et de catégoriser la capacité générale des masques à accumuler des bactéries lorsqu’elles sont utilisées par la population générale. Ceci également en ce qui concerne une évaluation des risques, en utilisant la pire considération qui est nécessaire dans une telle approche protectrice ( 47 ). En conséquence, nous avons entrepris une exploration microbiologique avec des échantillons aléatoires de masques faciaux utilisés par les membres de la population générale, ainsi qu’une revue systématique de la littérature rapide. Cette approche holistique combinée avec l’ARNr 16S (séquençage Sanger), la culture et l’analyse biochimique ainsi que la coloration rose au bengale plus une analyse de littérature systématique n’ont pas été réalisées auparavant et est la première du genre.
Matériels et méthodes
Coloration et visualisation du Bengale Rose de la contamination
La coloration avec du sel de sodium au bengale rose a été utilisée pour détecter la contamination des masques, comme décrit précédemment ( 45 ). La figure 1 illustre la zone du masque analysée.
Figure 1
Figure 1 . Coloration du Bengale Rose des masques de visage usés. La zone analysée est marquée par le cadre rouge. Les dimensions du masque indiquées par les fabricants (175 × 95 mm) excluent les plis, qui agrandissent la surface.
Conception d’étude de masque microbiologique
Dans cet échantillon pilote, des masques sur le visage chirurgical ont été collectés en mars 2022 (pendant l’obligation pandémique) de 15 volontaires consentants aléatoires (employés du département de l’université de Gdansk âgés de 19 à 65 ans), qui les avait portés pendant des périodes de 15 à 12 h h h. . Les détails du porteur n’ont pas été enregistrés car cela ne semble pas être crucial pour notre étude pilote, qui était destinée à montrer la contamination possible des masques utilisés par la population générale de travail. Cependant, avec notre échantillon aléatoire, nous avons capturé un profil d’utilisation réaliste avec des fluctuations temporelles typiques en raison des différents utilisateurs de la population générale. Chaque masque a été stocké dans un sac en plastique séparé jusqu’à l’examen. Les masques, à l’exclusion des boucles d’oreille, ont ensuite été coupés de manière aseptique en plusieurs pièces à l’aide de ciseaux stériles dans une armoire à débit laminaire. Ces pièces ont été transférées dans des tubes contenant 15 ml de solution saline tamponnée au phosphate stérile (PBS), équilibré pendant 1 min à température ambiante, puis vortexé pendant 30 s. Trois masques chirurgicaux inutilisés, propres et propres (Shandong Kaibo Medicinal Packaging Co., Ltd., Chine) ont été traités de manière identique comme des contrôles négatifs.
Pour déterminer le nombre de bactériens, les suspensions ont été diluées à 10 et 100 fois, puis des volumes de 100 μl ont été répartis sur une gélose Columbia contenant 5% de sang de mouton (Graso Biotech, Owidz, Pologne). Les plaques ont été incubées de manière aérobie pendant une nuit à 37 ° C, puis les colonies ont été comptées. La charge bactérienne a été déterminée comme des unités de formage de colonies par ml (CFU / ml) de suspension, puis rebelle comme CFU / masque ( 38 ). Dix colonies par masque usé ont été repensées, cultivées sur du bouillon de soja tryptique (Graso Biotech, Owidz, Pologne), puis stockée dans des solutions de stock de glycérol à 15% (v / v) à −70 ° C en attendant l’identification moléculaire.
Identification des isolats par séquençage Sanger du gène d’ARNr 16S
Quarante isolats ont été identifiés par PCR et séquençage Sanger du gène d’ARNr 16S. En bref, les colonies bactériennes ont été suspendues dans 30 μL d’eau stérile et lysées dans 95 ° C, suivies d’une centrifugation à 13 000 x g pendant 2 min. Les supernées ont été utilisées pour la PCR. Les amorces étaient: Forward F27 5′-AGAGTTTGATCMTGGCTCAG-3 ′ et inverse R1492 5′-CTACGGYTACCTTGTTACGACTT-3 ′ ( 48 , 49 ). Le mélange réactionnel (25 μL) contenait: 0,1 μm de chaque amorce, 1 μL de surnageant bactérien, 0,6 U de Taq polymérase (Eurx, Gdansk, Pologne), 0,2 mM DNTPS et Taq polymérase du tampon (EURX), contenant 15 mM de MGCL 2 . Les conditions de cycle impliquaient 94 ° C pendant 5 min; 30 cycles de 94 ° C pendant 1 min, 50 ° C pendant 1 min, 72 ° C pendant 1,5 min et une étape finale à 72 ° C pendant 5 min. Le séquençage de Sanger a été effectué chez Macrogen Europe (Amsterdam, Pays-Bas) sur un analyseur d’ADN 3730xl (ThermoFisher Scientific, Waltham, MA, USA). L’amplification par PCR était telle que décrite par Monciardini et al. ( 50 ). Les données de séquençage ont été analysées par FinchTV 1.4 (Geoshipa, Inc.; Seattle, WA, USA), 1 Les extrémités des lectures séquencées ont été coupées et les assemblages résultants ont été explosés dans la base de données NCBI. Les données de séquençage sont disponibles sur fighare à https://doi.org/10.6084/m9.figshare.23614797 (consulté le 2 juillet 2023).
Caractérisation biochimique des isolats
Tous les isolats séquencés ont été repensés sur une gélose sanguin de Columbia avec 5% de sang de mouton pour l’évaluation de l’hémolyse et sur une gélose au sel de mannitol (Graso Biotech, Owidz, Pologne) pour l’identification préliminaire de Staphylococcus spp. Les staphylocoques ont été testés en outre en utilisant le kit Staph Latex (Polex ™, Pro-Lab Diagnostics, Bromborough, Royaume-Uni) pour distinguer S. aureus des autres espèces.
Recherche de littérature systématique
Nous avons systématiquement recherché des études scientifiques évaluées par des pairs, jusqu’en juin 2023, qui ont analysé quantitativement la colonisation ou la contamination du tissu, du chirurgie, du N95 et des masques similaires par bactéries et champignons. La recherche a été effectuée à l’aide de PubMed et Medline et comprenait des évaluations qualitatives et quantitatives. Les termes de recherche ont été créés en fonction des critères définis dans le schéma PICO ( 51 ). Le terme de recherche non spécifique «masque» a été omis, car il comprend également des respirateurs et des masques de ventilation anesthésiologiques. Au lieu de cela, des termes spécifiques ont été choisis: «((masque facial) ou (facemask) ou (masque chirurgical) ou (FFP1) ou (FFP2) ou (FFP3) ou (N95) ou (KF94) ou (KN95)) et ((N95) contamination microbienne) ou (bactéries) ou (champignons)). » Deux chercheurs indépendants ont identifié et sélectionné des études éligibles. Les critères d’inclusion qualitatif étaient: présentation reproductible valide de la contamination microbienne, collection compréhensible de masques évalués, crédibilité des résultats et focalisation claire. Les critères d’inclusion quantitative étaient: des méthodes appropriées et précises, une mesure valide des résultats, une sélection représentative des masques évalués et des méthodes de détection / analyse reproductibles. Les articles sélectionnés ont été vérifiés par au moins trois des auteurs actuels pour l’admissibilité potentielle. La conception de l’étude, la méthodologie, les méthodes analytiques et expérimentales ont été évaluées. Les exclusions et les raisons ont été documentées. Pour les études incluses, les données suivantes ont été extraites dans des tableaux: auteur et année, méthode et type d’étude, taille de l’échantillon et type (s) de masque, du (s) du (s) des masques, des résultats / micro-organismes examinés, du contenu et des espèces principales. Des calculs et des graphiques mathématiques simples ont été effectués avec Libre-Office Calc, un package de bureau gratuit et open-source de la Fondation Document ( 52 ).
Résultats
Abondance et types de bactéries sur des masques usés
La contamination des masques usés était visible, macroscopiquement, après coloration au Bengale rose ( figure 2 ). Ce colorant se lie aux bactéries, aux champignons et aux cellules tissulaires ainsi qu’aux débris avec l’intensité de la couleur suggérée pour refléter le degré de contamination ( 53 – 57 ).
Figure 2
Figure 2 . Exemple masque la coloration avec la rose du Bengale, se liant aux cellules tissulaires, aux débris et aux bactéries.
Sur la base de la culture, la charge bactérienne moyenne de masque facial chirurgical propre et jamais utilisé était de 0,1 × 10 3 CFU a récupéré / masque tandis que la charge moyenne arithmétique sur les masques utilisés était de 4,24 × 10 4 CFU récupéré / masque (moyenne géométrique 1,3 × 10 4 ). Les bactéries étaient les plus abondantes sur les masques usés 5 et 6, avec 1,03 × 10 5 et 2,85 × 10 5 CFU a récupéré / masque, respectivement ( tableau 1 ). L’identification biochimique et moléculaire a révélé des espèces de staphylococciques sur ces deux derniers masques, notamment S. aureus , S. Warneri et S. epidermidis ( tableau supplémentaire 2 ). Bien que la morphologie des colonies différait entre les masques, les phénotypes dominants, dans presque tous les cas, y compris les masques inutilisés, étaient les petites colonies blanches typiques de S. epidermidis et d’autres staphylocoques négatifs de la coagulase ( figure supplémentaire 1 ).
Tableau 1
Tableau 1 . L’abondance de bactéries dans les masques.
Identification des isolats par séquençage Sanger du gène d’ARNr 16S
Sur 52 colonies soumises à la PCR, nous avons choisi les 40 avec l’amplification du produit la plus efficace pour le séquençage. Les résultats de l’explosion détaillés sont présentés dans le tableau supplémentaire 1 .
La grande majorité (32, 80%) de ces 40 appartenait au genre Staphylococcus confirmant les identifications phénotypiques. Nous avons identifié quatre espèces de coagulases négatives: S. epidermidis (la plus abondante), S. warneri , S. pasteuri et S. hominis , qui appartiennent toutes à la peau humaine normale et au microbiote nasal ( tableau supplémentaire 2 ) ( 58 ). de coagulase positif Sur le masque 5, nous avons confirmé Staphylococcus ( tableau supplémentaire 2 ) avec S. aureus et S. argenteus .
Quatre autres colonies séquencées comprenaient des espèces de Bacillus formant des endospores , à savoir B. cereus, B. Thuringiensis, B. altitudinis, B. megaterium et autres ( tableaux supplémentaires 1, 2 ), qui sont des bactéries du sol ( 59 ). Parmi les quatre colonies identifiées restantes (« autres » de la figure 3 ), nous avons trouvé Sporosarcina newyorkensis , une autre tige gram-positive formant des endospores, parfois récupérée des bactériémies humaines et du lait de vache ( 60 ). La seule espèce à Gram négatif trouvée était la pseudomonad psychrobacter faecalis ( tableau supplémentaire 2 ), une espèce psychrophile associée aux fèces de pigeons ( 61 ) et rapportées également à partir d’échantillons humains ( 62 ). Nous n’avons pas isolé les streptocoques, bien que ce soit une composante majeure du microbiote oral humain. Peut-être que leurs taux de survie sur les masques sont faibles, ou leur récupération nécessite une incubation enrichie de CO 2 , et non une incubation d’air telle qu’elle est utilisée ici.
Figure 3
Figure 3 . L’abondance relative de différentes espèces bactériennes a été récupérée de masques.
Identification biochimique des isolats
Les mêmes 40 colonies ont été soumises à une identification biochimique, ce qui donne des résultats cohérents avec le séquençage. L’hémolyse a été détectée pour presque toutes ces bactéries ( tableau supplémentaire 2 ) bien que son intensité soit très variable ( tableau supplémentaire 2 ; figure supplémentaire 1 ). La plupart des bactéries ont montré une halotolérance mais seulement cinq mannitol fermentés: ces derniers ont été testés pour la coagulase et la protéine A et trois, tous à partir du masque 5, se sont avérés positifs pour les deux caractères, confirmant l’identification comme S. aureus ( tableau supplémentaire 2 ); Tous avaient une morphologie typique de l’espèce ( figure supplémentaire 1 , masque 5).
Recherche de littérature systématique
La recherche documentaire a initialement donné 1 310 résultats. Cela a été rétréci (voir le diagramme PRISMA, figure 4 ) à 14 études évaluant la contamination bactérienne et fongique des masques de tissu, chirurgicaux et N95, portés pour des périodes allant de 5 min à 3 jours. Onze études ont considéré les bactéries, cinq champignons et trois ( tableau 2 ). Quatre études étaient destinées à la population générale, tandis que 10 étaient pour les travailleurs de la santé (HCWS) ( 38 , 41 , 42 , 44 , 46 , 63 – 71 ). Six étaient destinés aux unités chirurgicales (une procédés spécifiquement de la chirurgie orthopédique) et cinq pour les pratiques dentaires ( 44 , 64 – 67 ). Seuls deux ont fourni une quantification exacte et une identification bactérienne par l’ARNr 16S; Ceux-ci ont tous deux étudié la population générale ( 38 , 63 ). Les résultats de la recherche documentaire sont résumés dans l’extraction ( tableau 2 ).
Figure 4
Figure 4 . PRISMA FLUX THART pour la recherche de littérature.
Tableau 2
Tableau 2 . Résultats microbiologiques de la recherche documentaire (contamination du masque par des bactéries et des champignons).
Discussion
Nous avons trouvé une forte contamination bactérienne des masques chirurgicaux portés par la population générale, avec jusqu’à 2,85 × 10 5 CFU / masque (moyenne 4,24 × 10 4 ).
Malheureusement, il n’y a pas de normes microbiologiques pour les masques usés contre lesquels examiner ces résultats; Dans l’UE, la seule exigence de bioburden pertinente est en 14683 pour les nouveaux masques, nécessitant ≤ 30 CFU / g. Néanmoins, comme les masques représentent un système de filtrage en amont des voies respiratoires, les valeurs limites des systèmes de ventilation sont pertinentes, notamment la norme allemande pour les surfaces de ventilation et de climatisation, VDI 6022, partie 4 ( 72 ). Cela spécifie des dénombrements de 25 à 100 CFU / 25 cm 2 comme «limite», tandis que les surfaces avec des comptes> 100 CFU / 25 cm 2 nécessitent une action ou un remplacement immédiat.
Un masque chirurgical jetable a une surface unique de CA. 230 cm 2 ( 73 ), ce qui signifie que dans notre pire cas (2,85 × 10 5 CFU / masque = 3,09 × 10 4 CFU / 25 cm 2 ), la limite supérieure de VDI 6022 a été dépassée par ca. 310 fois (moyen 46 fois) ( tableau 1 ). Les valeurs d’une étude comparable montrent un dépassement de 166 fois avec des masques de coton ( 38 ); Une autre étude, pour les agents de santé avec des masques chirurgicaux portés pour une période non spécifiée, a indiqué> 2 000 fois dépassement ( tableau 2 ) ( 44 ). Il convient d’ajouter que la charge bactérienne d’un masque se trouve directement devant les voies respiratoires alors que l’évent d’un système de climatisation se trouve généralement à plusieurs mètres.
Les exigences EN 14683 pour les nouveaux masques ont également été largement dépassées pour les articles usés ( tableau 2 ), sur la base des poids de ca. 3 g pour un masque chirurgical et 4 g pour les masques N95 / FFP2 ( 74 ); Le dépassement de cette exigence était évident même pour les masques non portés ( tableau 1 ).
La lourde contamination générale des masques usés était encore démontrable par coloration au bengale rose ( figure 2 ).
L’étude microbiologique du masque utilisée a été principalement des staphylocoques cutanés à la coagulase et des bactéries du sol formant des endospores ( Bacillus spp.) Sur utilisée ( figure 3 ). Cette prédominance des staphylocoques est conforme à d’autres études sur les masques de visage contaminés dans la population générale et les travailleurs de la santé ( 42 , 44 , 64 – 66 , 68 ). Un masque (n ° 5) a été contaminé par S. aureus , un agent pathogène bien connu et polyvalent ( figure 3 ; tableau 1 ) ( 75 – 78 ). Jusqu’à 30% de la population transporte S. aureus nasal sans symptômes ( 79 ), mais avec un risque accru d’auto-infection ( 75 ). La contamination contingente des masques peut faciliter la diffusion de S. aureus et, une infection cutanée plausiblement ( 75 ). Une association entre le transport nasal et la contamination chirurgicale et chirurgicale ainsi que le masque KN95 a été montrée précédemment pour S. aureus et même pour les non-porteurs, l’organisme a été fréquemment détecté sur les masques KN95 ( P = 0,04, test exact de Fisher) impliquant des sources exogènes de contamination (mains, environnement et gouttelettes externes contenant des flux d’air, etc.) ( 75 ). À l’appui de cela, certains auteurs notent que S. aureus contamine sur les surfaces externes et internes des masques ( 75 ).
Plusieurs auteurs ont associé l’utilisation des éruptions cutanées masques du visage, certaines impliquant S. aureus ( 80 ), y compris une nouvelle occurrence ou une exacerbation de l’acné, de la rosacée et de la dermatite séborrhoéique ( 81 ). D’autres auteurs notent l’enrichissement du microbiote des yeux normaux avec S. aureus de la respiration et des gouttelettes expirés tout en portant un masque contribuant au développement de l’inflammation des paupières (Chalazion) ( 82 , 83 ) et des infections de la cornée ( 84 ), également des infections pour les yeux plus profonds Dans le contexte des traitements (endophtalmiste après vitrectomie) ( 85 ). Il existe également des preuves que S. aureus peut augmenter la réplication du virus SARS-COV-2 de 10 à 15 fois ( 86 ), bien que cela semble plus pertinent dans le nez supérieur que sur un masque, où le virus est peu probable être reproduit.
Parmi d’autres staphylocoques, nous avons trouvé principalement S. epidermidis ( figure 3 ). D’une part, c’est un composant normal et inoffensif du microbiote cutané; De l’autre, cela peut être un danger pour les individus vulnérables immunodéprimés ( 87 – 89 ). Même chez les individus en bonne santé, les staphylocoques coagulases négatives, à grande abondance, peuvent contribuer à des affections cutanées inflammatoires telles que la dermatite atopique et l’acné vulgaris ( 58 , 90 – 92 ) avec des preuves que le port d’un masque a considérablement augmenté l’incidence de l’acné en particulier ( 93 – 101 ).
Nous avons également trouvé Bacillus spp. dans les masques, y compris les espèces qui produisent des entérotoxines ( 59 ). Bien que la croissance bactérienne des masques puisse être possible (voir ci-dessous), nous n’avons vu aucune preuve que la croissance a atteint les niveaux – typiquement> 10 6 / G – associé aux toxines dans les aliments ( 102 ). De plus, les porteurs (sauf peut-être les enfants) sont peu susceptibles de mâcher leurs masques, ce qui signifie que ces organismes peuvent être rejetés comme un risque.
Revue de la littérature sur la contamination du masque
Notre revue de la littérature a montré que tous les types de masques pertinents (chirurgicale, N95, tissu) deviennent de plus en plus contaminés par des micro-organismes pendant l’usure ( tableau 2 ; figure 5 ) ( 38 , 40 , 41 , 46 , 65 , 67 , 71 ).
Figure 5
Figure 5 . Dépendance temporelle de la contamination du masque facial pendant l’usure, en fonction des données de la littérature ( tableau 2 ). Les diagrammes indiquent l’association entre le CFU / masque et la durée d’usure, sur la base des valeurs moyennes de trois publications ( 41 , 46 , 67 ). S’ils sont inclus dans les études primaires, les écarts-types sont également montrés. Yang et al. a étudié les surfaces intérieures des masques portés par la population générale, tandis que Liu et al. et Checkchi et al. a examiné les couches extérieures de masques portés par HCW.
The literature reports contamination by bacteria of the genera Acinetobacter, Bacillus, Escherichia (specifically, E. coli , a faecal organism), Enterobacter, Enterococcus (another faecal organism), Klebsiella (including K. pneumoniae ), Micrococcus, Pseudomonas, Staphylococcus (including S. aureus ) et Streptococcus et par des champignons des genres Aspergillus, Alternaria, Candida, Cadosporium, Microsporum et Mucor ( tableau 2 ). Ces organismes sont nourris par la salive humaine, le biofilm oral nébulisé et les condensats de la respiration expirés, créant une préoccupation de biosécurité sous-estimée.
Dans la population générale, la contamination du masque interne dépasse généralement l’extérieur pour les bactéries – et peut-être, bien que cela varie selon l’étude – en outre pour les champignons ( tableau 2 ) ( 63 , 70 ). Pour les travailleurs de la santé utilisant des masques chirurgicaux, en revanche, la contamination externe dépasse la contamination interne à la fois pour les bactéries et les champignons ( p <0,001) ( 42 , 44 , 64 ) et corrélé avec la qualité de l’air microbiologique dans les domaines où ces employés travaillaient ( 42 ). Pour les masques N95, cependant, la contamination bactérienne interne apparaît plus élevée que dans les établissements de soins de santé ( 68 ). De plus, la contamination bactérienne totale des masques N95 usés a dépassé celui des masques chirurgicaux usés de manière similaire ( 68 ).
La contamination fongique est observée jusqu’à 70 à 88% des masques utilisés ( 70 , 71 ), et peut également être plus élevé à l’intérieur de l’extérieur du masque ( 70 ). Cela est surprenant, étant donné que les champignons doivent provenir de l’extérieur du masque ( 63 ).
Une comparaison du nombre de masques faciaux bactériens maximaux pour les travailleurs de la santé et la population générale, en fonction des données du tableau 2 et des temps de port / utilisation entre 5 min et 3 jours, ont montré une grande variance des données en raison de la variance des temps de port et des utilisateurs et environnementaux Facteurs. Il y a une tendance à des charges bactériennes plus élevées dans la population générale ( tableau 2 ). Ces résultats peuvent refléter une utilisation plus large et prolongée dans la population générale ( 7 , 8 ). En raison du petit nombre d’études similaires, une évaluation statistique méta-analytique n’a pas été effectuée.
Contamination du masque en face – facteurs de contribution
Les masques sont une bonne matrice pour l’accumulation microbienne et, potentiellement, la croissance, la conservation d’une température supérieure à ambient ( 103 – 107 ), l’humidité et les débris riches en nutriments ( 38 – 41 , 45 , 108 ). Outre les substances aspirées de l’extérieur, les nutriments comprennent des protéines expirées et autres débris, des cellules épithéliales exfoliées et mortes. Les gouttelettes de condensation dans le souffle expiré contiennent des métabolites, des sels, des lipides et des protéines non volatils ainsi que des bactéries et des virus intacts et dégradés ( 109 ). Cette richesse organique a été visualisée dans notre coloration aux roses du Bengale. La croissance, plutôt que la simple survie ( 38 , 39 , 41 , 45 , 108 , 110 ) des colonies bactériennes et fongiques est révélée par microscopie électronique à balayage des masques faciaux (FFP2) usée pendant plusieurs heures ( 40 ).
L’espace mort des masques N95 rigides offre un environnement humide et humide ( 103 ) avec une humidité relative de 1,5 à 2,6 fois plus élevée que l’extérieur ( 41 ) augmentant à 100% après 60 min d’utilisation ( 40 ). Cela peut créer un terrain reproducteur particulièrement attrayant pour les bactéries ( 41 ) expliquant les résultats (ci-dessus) que les masques N95 deviennent plus fortement contaminés que les masques chirurgicaux et que, dans les soins de santé, la contamination interne a dépassé externe, inversant le modèle observé pour les masques chirurgicaux ( 68 ).
Les micro-organismes piégés et incubés dans le masque peuvent être distribués au porteur, à l’environnement et à d’autres ( 16 , 111 – 113 ). Si la fuite, en raison d’un défaut ou d’un mauvais ajustement, affecte 1% de la zone du masque, l’efficacité de filtration est réduite de 50%; Si l’écart est de 2% de la zone du masque, l’efficacité est réduite de 75% ( 114 ). De plus, l’efficacité de filtration d’exhalation est significativement inférieure à l’efficacité de filtration théorique – étant respectivement de 12,4 et 46,3% pour les masques chirurgicaux et N95 ( 115 ). Dans les salles d’opération, la durée de port recommandée est limitée à quelques heures ( 116 ) car les masques chirurgicaux perdent leur efficacité au fil du temps ( 117 ). Alors qu’un masque frais a presque complètement empêché la contamination bactérienne d’une plaque d’agar maintenue à 10 à 12 cm de la bouche, cette efficacité a été mesurablement réduite dans les 30 minutes et négligeable après 2 h ( 118 ). Cette brève période d’efficacité de filtration a été encore réduite si le masque était mal ajusté ( 114 , 119 ) ou mouillé ( 119 ).
La pénétration des micro-organismes entre les couches du masque est possible, par l’action capillaire en fonction de l’humidité et des organismes spécifiques entre autres facteurs ( 120 ). À son tour, peut faciliter la formation de minuscules gouttelettes chargées d’organisme. Ceux-ci peuvent ensuite être projetés ou inhalés à chaque respiration ( 16 , 111 , 114 , 115 , 121 – 123 ). Dans ce contexte, nous soulignons la respiration à prédominance orale tout en portant un masque ( 16 , 124 ), contrairement à la respiration normale sans entrave, qui est largement via le nez, avec une plus grande filtration. La respiration orale augmente le danger d’inhaler directement les micro-organismes du masque dans les voies respiratoires plus profondes ( 125 ). Dans une étude humaine avec un aérosol radiomarqué et des diamètres de particules moyens de 4,4 μm (plage de 3,8 à 5,1 μm) ont trouvé une forte augmentation du dépôt dans les poumons (+ 37%) lors de la respiration par rapport à via le nez (75% Vs. 38%) ( 126 ). De plus, les masques – et en particulier le type N95 – la clairance muciliaire naturelle des voies respiratoires supérieures, améliorant davantage l’inhalation et la distribution des bactéries ( 127 ).
Enfin, dans le contexte, les masques faciaux contiennent des plastiques, auxquels les micro-organismes peuvent s’adsorber ( 40 , 128 ). Par conséquent, ainsi que des aérosols, les micro-particules en plastique peuvent également être libérées par des masques ( , 129-133 ) agissant comme porteurs pour la distribution des bactéries pathogènes et des champignons ( 134 ). Fait intéressant, il n’y a pratiquement pas de surface ou de matériau, pas même la peau nue, qui garantit une telle survie et une préservation à long terme de l’infectivité pour les virus comme le réseau plastique-polypropylène des masques, dans lequel les virus SARS-COV-2 sont stockés et restent contagieux jusqu’à 2 semaines, même lorsqu’ils sont séchés ( 135 ).
Contamination du masque de visage – implications cliniques potentielles
Dans une étude transversale pré-cuve sur 710 individus, le port (pour des raisons religieuses) de revêtements faciaux en tissu par les femmes saoudiennes, tirés de la population générale, a été associé à des incidents statistiquement accrus de «rhume» et d’asthme (17) ( 17 ) . Ailleurs, les changements de peau physiopathologique ( 136 ) étaient associés à la port de masques dans la population générale et les travailleurs de la santé ( 137 , 138 ). Plusieurs auteurs ont trouvé des changements dans la métabolomique cutanée, avec un risque accru de perturbation et d’inflammation des barrières, en raison de dysbioses du microbiome cutané ( 136 , 139 , 140 ) conduisant – ou favorisant le développement de la dermatite atopique et de l’acné vulgaris ( 139 ). Dans le contexte, les respirateurs N95 ont provoqué un trouble plus important que les masques chirurgicaux ( 139 ).
Les conditions oculaires ont également été associées à l’utilisation du masque ( 82 – 85 , 121 , 141 – 145 ), tandis qu’Islam et al. ont trouvé des preuves indirectes de changements dans le microbiome oral ( 146 ). Sukul et al. Changements dans le microbiome intestinal (altérations métaboliques) ( 19 ) tandis que Xiang et al. a trouvé le changement des communautés microbiennes nasales après un usage de masque prolongé ( 110 ). Enfin, les masques de visage sont mentionnés comme facteurs possibles derrière une augmentation des cas de mucormycose pendant la pandémie Covid-19, en particulier chez les individus immunodéprimés ou autrement vulnérables ( 70 , 71 , 147 ).
Pratiques pour minimiser la contamination microbienne
Il y a des considérations générales pour l’utilisation de masques faciaux dans n’importe quelle situation, ainsi que des conseils officiels sur leur utilisation appropriée ( 16 , 129 , 148 ). La minimisation de la contamination microbienne est essentielle pour assurer leur utilisation sûre, en particulier dans les soins de santé. L’OMS recommande d’éviter de toucher la surface du masque, également que les masques doivent être stockés dans un endroit propre et sec des contaminants potentiels ( 6 ). Les masques jetables doivent être supprimés après chaque utilisation et non réutilisés. La formation doit être dispensée sur la façon de mettre et de retirer des masques afin d’éviter la propagation microbienne et l’auto-infection. L’OMS recommande en outre de nettoyer les mains avant de toucher un masque (avant et après le retirer). Lorsque le masque est retiré, il doit être stocké dans un sac en plastique propre ou éliminé dans une poubelle de déchets ( 6 ).
Dans certaines situations, un bouclier facial peut être utilisé conjointement avec des masques pour fournir une barrière supplémentaire contre la contamination. Enfin, le masque doit être porté aussi court que possible, non seulement pour des raisons microbiologiques (contamination dépendante du temps du masque facial pendant le port), mais aussi pour des raisons toxicologiques et physio-métaboliques ( 14 , 129 ).
Il est évident que de grandes sections de la population, y compris les enfants, ne sont pas en mesure de suivre ces instructions complexes de manière adéquate et cohérente ( 148 ). Les alternatives aux masques doivent être recherchées et priorisées (par exemple, les systèmes de ventilation, les mesures d’hygiène et autres).
Résultats en contexte
Bien avant la pandémie, les masques faciaux sont devenus largement utilisés dans les soins de santé en médecine (notamment la chirurgie) et dans certaines industries manufacturières ( 16 , 149 – 151 ), visant à prévenir ou à minimiser l’infection ou la contamination ( 8 , 14 , 73 , 151 – 159 ). Néanmoins, leur efficacité dans les milieux de santé a été discutable bien avant 2020 ( 160 ) et leur rôle dans l’opération reste controversé ( 161 ). Compte tenu de cette histoire, il y a eu étonnamment peu de recherches sur les effets de l’utilisation à long terme des groupes professionnels. Bien que les masques filtrent les débris plus grands et les gouttelettes d’aérosols de l’air, ils comportent les risques microbiologiques décrits ici avec des dommages toxicologiques, physiologiques, psychologiques et sociologiques ( 14 , 16 , 18 – 35 , 129 , 162 ).
Les risques et les avantages d’exiger l’utilisation du masque par les populations doivent être pesés par des points de vue éthiques et médicaux ( 13 , 14 , 16 , 163 , 164 ). Pour que les masques soient exigés, les effets secondaires et les risques doivent être inférieurs au risque de ne pas porter de masque. Une évaluation de Cochrane standard en or, basée sur des essais cliniques ( 12 ) n’a trouvé aucune preuve substantielle d’efficacité dans la prévention des infections respiratoires virales et une étude récente, bien qu’avec plusieurs facteurs de confusion possible -19 Infection ( 165 ). D’un autre côté, les dommages potentiels sont nombreux ( 2 , 3 , 5 , 14 – 16 , 19 – 23 , 36 , 37 , 166 – 172 ). Ils comprennent les MIES ( 16 ), les altérations nocives de gaz sanguin ( 14 , 19 ) et les risques microbiologiques potentiels décrits ici. Les masques ne doivent pas être obligatoires pour la population générale compte tenu de cet équilibre des preuves contre leur utilisation. Ces points ont été soulevés par de nombreux scientifiques ( 14 , 16 , 17 , 36 , 37 , 129 , 166 , 173 – 175 ), y compris les principaux experts en respiration ( 176 ).
Limitations et forces
Les forces de notre article sont l’utilisation d’une méthode précise – séquençage d’ARNr 16S – pour identifier les bactéries trouvées. De plus, nous avons entrepris un aperçu de la littérature systématique et discutons des résultats des perspectives microbiologiques et cliniques holistiques. Les masques collectés dans notre étude ont été fournis par des individus aléatoires au cours de la vie quotidienne, représentant un échantillon de population générale réaliste. Coloration au Bengale de la rose visualisée de manière frappante une contamination approfondie. À la fois, notre taille limitée de l’échantillon et notre revue de littérature rapide ne doivent être considérées que comme une évaluation pilote, avec une analyse plus approfondie nécessaire. En raison du petit nombre d’études de la même conception, une méta-analyse n’a pas été réalisée. La force de cette revue est plutôt qualitative, cataloguant la vaste littérature scientifique publiée par de nombreux scientifiques dans le monde sur plusieurs décennies, démontrant des preuves expérimentales de la contamination du masque facial et de ses risques.
Conclusion
Notre étude expérimentale et la littérature publiée montrent que les masques faciaux accumulent des micro-organismes, y compris les pathobiontes ( tableaux 1 , 2 ) ( 38 , 41 , 42 , 44 , 46 , 63 – 71 , 177 ), avec une charge microbienne jusqu’à plusieurs centaines de fois supérieur à la limite de VDI 6022 standard allemande pour les surfaces des systèmes de ventilation ( 72 ) et les exigences EN 14683 pour les masques inutilisés. La contamination augmente avec le temps de port prolongé ( figure 5 ) ( 38 , 41 , 46 , 65 – 67 , 70 , 71 ) et est plus élevé pour N95 que les masques chirurgicaux ( 68 ). La plupart des contamination ont été avec des staphylocoques, y compris parfois le pathogène S. aureus .
En termes simples: (i) Le masque agit comme un piège à filtre avec des bactéries s’accumulant sur ses surfaces externes et internes; (ii) le masque agit ensuite comme un «incubateur microbiologique» à l’entrée des voies respiratoires; (iii) Les micro-organismes peuvent se développer dans le masque, nourris par des débris cutanés, du mucus et du «condensat expiré» ( 16 , 38 , 39 , 41 , 45 , 108 – 110 ). Ces organismes / agents pathogènes piégés peuvent alors être inhalés, favorisant l’infection des voies respiratoires ( 17 , 37 ) ou, lorsqu’ils sont distribués via des flux d’air ( 111 , 114 , 115 , 122 , 142 , 143 , 178 , 179 ) l’œil ( 82 – 85 , 121 , 142 ). De plus, le microbiome cutané est perturbé, conduisant ou promouvant d’autres infections et conditions allergiques ( 38 , 77 , 110 , 140 , 180 ).
Enfin, les micro-organismes accumulés peuvent être distribués par fuite ( 111 , 114 , 115 ), amplifiés par l’effet d’atomiser du masque ( 14 , 16 , 122 , 181 , 182 ).
Une analyse de Cochrane, basée uniquement sur le plus haut niveau de preuve, n’a trouvé aucune preuve que les masques réduisaient la propagation des infections virales respiratoires dans la population générale ( 12 ). D’un autre côté, leurs préjudices, au-delà des personnes étudiées ici, sont claires. Ils entravent la communication ( 32 – 34 , 94 , 183 – 188 ). Ils entravent l’apprentissage, en particulier pour les enfants ( 2 , 3 , 5 , 14 , 26 , 35 , 148 , 162 , 171 , 174 , 177 , 189 ). Ils sont associés à une hypoxémie transitoire (diminution du sang O 2 ), hypercarbia transitoire (augmentation du sang CO 2 ) ( 14 , 16 , 19 , 21 – 23 , 171 , 172 ). Ils nient le porteur de l’individualité la plus fondamentale – de montrer leur visage ( 26 , 27 , 30 – 34 , 162 , 189 ). Leur imposition à long terme est particulièrement nocive pour les membres vulnérables de la population ( 14 , 16 , 19 ). Des articles scientifiques récents indiquent des problèmes toxicologiques via l’inhalation de particules plastiques et des composés organiques cancéreux provenant du matériau du masque ( 14 , 18 , 129 , 133 ).
En bref, les effets indésirables des masques sont clairs ( 2 , 3 , 5 , 16 , 18 , 19 , 23 , 36 , 129 , 166 – 172 , 190 ), tandis que l’effet antiviral protecteur dans les scénarios réel reste douteux ( 12 – 15 , 165 , 175 , 191 – 209 ). Compte tenu de cela, ainsi que les problèmes de contamination microbiologique mis en évidence, les lois et les exigences de masquage ne répondent pas à l’éthique médicale de base de «ne pas de mal». Les lois et les mandats exigeant une utilisation du masque en conséquence n’ont pas de place valable dans la gestion pandémique respiratoire.
Énoncé de disponibilité des données
Les ensembles de données présentés dans cette étude se trouvent dans les référentiels en ligne. Les noms du référentiel / des référentiels et des numéros d’adhésion se trouvent dans l’article / matériel supplémentaire .
Contributions des auteurs
KK: Conceptualisation, conservation des données, analyse formelle, acquisition de financement, enquête, méthodologie, administration de projet, ressources, logiciels, supervision, validation, visualisation, écriture – brouillon original, écriture – revue et montage. BW: Conceptualisation, conservation des données, analyse formelle, enquête, acquisition de financement, méthodologie, ressources, logiciels, supervision, validation, visualisation, écriture – brouillon original, écriture – revue et montage. AZ: Curration des données, enquête, visualisation, écriture – brouillon original, rédaction – revue et montage. DL: Analyse formelle, enquête, supervision, validation, rédaction – Braft original, rédaction – revue et montage. AJ-K: Conceptualisation, conservation des données, analyse formelle, acquisition du financement, enquête, méthodologie, administration de projet, ressources, logiciels, supervision, validation, visualisation, écriture – brouillon original, rédaction – revue et montage.
Financement
Les auteurs déclarent que le soutien financier a été reçu pour la recherche, la paternité et / ou la publication de cet article. La publication de cet article a été partiellement financée par l’Université de Gdansk.
Remerciements
Nous tenons à remercier le Dr Jadwiga Gronczewska du Département de génétique évolutive et de biosystème de l’Université de Gdańsk, en Pologne, pour son précieux soutien technique sur ce projet. Nous remercions également le Dr Bermpohl, hygiéniste et microbiologiste, et l’ophtalmologiste et physicien Mphys, le Dr Mengedoht (Gütersloh, Allemagne), qui ont inspiré des parties du manuscrit.
Conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de relations commerciales ou financières qui pourraient être interprétées comme un conflit d’intérêts potentiel.
Note de l’éditeur
Toutes les réclamations exprimées dans cet article sont uniquement celles des auteurs et ne représentent pas nécessairement celles de leurs organisations affiliées, ou celles de l’éditeur, des éditeurs et des examinateurs. Tout produit qui peut être évalué dans cet article, ou réclamation qui peut être fait par son fabricant, n’est pas garanti ou approuvé par l’éditeur.
3. Thomson, S. Mask mandater les enfants pendant la pandémie Covid-19: une perspective internationale des droits de l’homme. Scand J Public Health . (2022) 50: 683–5. doi: 10.1177 / 14034948221081087
4. Organisation mondiale de la santé, Fonds UNC (UNICEF). Who – Conseils sur l’utilisation des masques pour les enfants de la communauté dans le contexte de Covid-19: Annex aux conseils sur l’utilisation des masques dans le contexte de Covid-19, 21 août 2020. (2020). Disponible sur: https://apps.who.int/iris/handle/10665/333919 (consulté le 7 novembre 2020).
5. Schwarz, S, Jetzky, E, Krafft, H, Maurer, T et Martin, D. Corona Enfant Studies «Co-Ki»: premiers résultats d’un registre allemande sur la bouche et le nez couvrant (masque) chez les enfants chez les enfants . Monatschr Kinderheilkd . (2021) 169: 353–65. doi: 10.1007 / s00112-021-01133-9
6. Organisation mondiale de la santé. Who – Conseils sur l’utilisation des masques dans le contexte de Covid-19: Guide provisoire, 5 juin 2020. (2020). Disponible sur: https://apps.who.int/iris/handle/10665/332293 (consulté le 7 novembre 2020).
8. Gralton, J et McLaws, ML. Protéger les agents de santé contre la grippe pandémique: N95 ou masques chirurgicaux? Crit Care Med . (2010) 38: 657–67. doi: 10.1097 / ccm.0b013e3181b9e8b3
10. Roberge, R. Facemask Utilisation par les enfants lors d’épidémies infectieuses des maladies. Biosecur Bioterror . (2011) 9: 225–31. doi: 10.1089 / bsp.2011.0009
11. Munro, APS et Hughes, RC. Les couvrages du visage ont peu d’utilité pour les jeunes enfants d’âge scolaire. Arch Dis Child . (2023) 108: 77–8. doi: 10.1136 / archdischild-2022-324809
14. Kisielinski, K, Wagner, S, Hirsch, O, Klosterhalfen, B et Prescher, A. Toxicité possible de l’exposition chronique au dioxyde de carbone associée à l’utilisation du masque facial, en particulier chez les femmes enceintes, les enfants et les adolescents – un examen de la portée. Heliyon . (2023) 9: E14117. doi: 10.1016 / j.heliyon.2023.e14117
15. Coma, E, Català, M, Méndez-Boo, L, Alonso, S, Hermosilla, E, Alvarez-Lacalle, E, et al. Décroisser le rôle de l’utilisation obligatoire du visage couvrant les masques pour le contrôle du SARS-COV-2 dans les écoles: une étude quasi-expérimentale imbriquée dans une cohorte basée sur la population en Catalogne (Espagne). Arch Dis Child . (2022) 108: 131–6. doi: 10.1136 / archdischild-2022-324172
16. Kisielinski, K, Giboni, P, Prescher, A, Klosterhalfen, B, Graessel, D, Funken, S, et al. Un masque qui couvre la bouche et le nez est-il exempt d’effets secondaires indésirables dans une utilisation quotidienne et exempts de dangers potentiels? Int J Environ Res Santé publique . (2021) 18: 4344. doi: 10.3390 / ijerph18084344
17. Ahmad, Efem, Mohammed, M, Al Rayes, AA, Al Qahtani, A, Elzubier, Ag et Suliman, Fae. L’effet du port du voile par les dames saoudiennes sur la survenue de maladies respiratoires. J asthme . (2001) 38: 423–6. doi: 10.1081 / jas-100001497
18. Ryu, H, et Kim, YH. Mesurer la quantité de composés organiques volatils nocifs inhalés à travers des masques. Ecotoxicol Environ Saf . (2023) 256: 114915. doi: 10.1016 / j.ecoenv.2023.114915
19. Sukul, P, Bartels, J, Fuchs, P, Trefz, P, Remy, R, Rührmund, L, et al. Effets des masques de visage protecteurs Covid-19 et des durées sur la physiologie hémodynamique respiratoire et les constituants de la respiration expirés. Eur respir j . (2022) 60: 2200009. doi: 10.1183 / 13993003.00009-2022
20. Al-Allaff, RGM, Al-Taee, Smy et Baker, Std. Quelques impacts immunologiques de l’utilisation du masque facial pendant la pandémie Covid-19. Pak J Biol Sci . (2021) 24: 920–7. doi: 10.3923 / pjbs.2021.920.927
21. Vakharia, RJ, Jani, I, Yadav, S et Kurian, T. pour étudier les changements aigus dans l’oxygénation du cerveau sur l’IRM chez les travailleurs de la santé à l’aide de kits de masque N95 et d’EPI pendant six heures par jour. Imagerie indienne J Radiol . (2021) 31: 893–900. doi: 10.1055 / s-0041-1741086
22. Law, CSW, LAN, PS et Glover, GH. Effet du port d’un masque facial sur le contraste en gras IRMf. Neuroimage . (2021) 229: 117752. doi: 10.1016 / j.neuroimage.2021.117752
23. Patel, S, Mohapatra, E, Suntanthy, AK, Shah, S, Abraham, J, Nanda, R, et al. Une étude pilote pour évaluer les changements dans les paramètres du gaz sanguin veineux et les biomarqueurs de l’hypoxie chez les travailleurs de la santé à l’aide de différents types de masques. Inde pulmonaire . (2023) 40: 134–42. doi: 10.4103 / Lungindia.Lungindia_343_22
24. Prousa, D. Étude sur les plaintes psychologiques et psychovegetatives avec les réglementations actuelles de protection du nez buccal. Psycharchives . (2020) 1–128. Doi: 10.23668 / psycharchives.
25. Pavlova, MA, Carbon, CC, Coello, Y, Sokolov, AA et Proverbio, Am. Éditorial: Impact de la couverture du visage sur la cognition sociale et l’interaction. Neurosci avant . (2023) 17: 1150604. doi: 10.3389 / fnins.2023.1150604
26. Carbon, CC, Held, MJ et Schütz, A. La lecture des émotions dans les visages avec et sans masques est relativement indépendante de l’exposition étendue et des variables de différence individuelles. Front Psychol . (2022) 13: 856971. doi: 10.3389 / fpsyg.2022.856971
28. Villani, C, D’Ascenzo, S, Scerrati, E, Ricciardelli, P, Nicoletti, R et Lugli, L. Le port du masque facial affecte notre attention sociale sur l’espace. Front Psychol . (2022) 13: 923558. doi: 10.3389 / fpsyg.2022.923558
29. Proverbio, AM et Cerri, A. La reconnaissance des expressions faciales sous masques chirurgicaux: la primauté de la colère. Neurosci avant . (2022) 16: 864490. doi: 10.3389 / fnins.2022.864490
30. Grundmann, F, Epstude, K et Scheibe, S. Les masques de visage réduisent la précision de la reconnaissance des émotions et la proximité perçue. Plos un . (2021) 16: E0249792. doi: 10.1371 / journal.pone.0249792
31. Mathis, L. Les effets des masques du visage sur l’interprétation des émotions chez les individus socialement anxieux. Grad Stud J Psychol . (2023) 20: 88–98. doi: 10.52214 / gsjp.v20i1.10167
32. Truong, TL, Beck, SD et Weber, A. L’impact des masques faciaux sur le rappel des phrases parlées. J ACOULT SOC AM . (2021) 149: 142–4. doi: 10.1121 / 10.0002951
33. Sönnichsen, R, Llorach Tó, G, Hochmuth, S, Hohmann, V et Radeloff, A. Comment les masques du visage interfèrent avec la compréhension de la parole des individus à halence normale: la vision fait la différence. Otol neurotol . (2022) 43: 282–8. doi: 10.1097 / mao.000000000000003458
34. McKenna, VS, Kendall, CL, Patel, Th, Howell, RJ et Gustin, RL. Impact des masques faciaux sur l’acoustique de la parole et l’effort vocal chez les professionnels de la santé. Laryngoscope . (2022) 132: 391–7. doi: 10.1002 / lary.29763
37. Fögen, Z. L’effet Foegen: un mécanisme par lequel les masques facilitantes contribuent au taux de mortalité de la cas Covid-19. Médecine (Baltimore) . (2022) 101: E28924. doi: 10.1097 / md.0000000000028924
38. Delanghe, L, Cauwenberghs, E, Spacova, I, de Boeck, I, Van Beeck, W, Pepermans, K, et al. Masques faciaux en coton et chirurgical en milieu communautaire: contamination bactérienne et hygiène du masque facial. Front Med (Lausanne) . (2021) 8: 732047. doi: 10.3389 / fmed.2021.732047
40. Buzzin, A, Domenech-Gil, G, Fraschecti, E, John, E, Pupils, D et Chapter, D. Assexing Le consécutif de l’utilisation prolongée de l’utilisation prolongée de jeturs à l’aide de massages. SCIS Rep . (2022) 12: 16796. Deux: 10.1038 / S41598-022-022-20692-9
41. Yang, Q, Li, H, Shen, S, Zhang, G, Huang, R, Feng, Y, et al. Étude de la distribution micro-climat et bactérienne dans l’espace mort des respirateurs de la face de filtrage N95. Sci Rep . (2018) 8: 17382. doi: 10.1038 / s41598-018-35693-W
42. Luksamijarulkul, P, Aiempradit, N, et Vatanasomboon, P. Contamination microbienne sur les masques chirurgicaux utilisés parmi le personnel de l’hôpital et la qualité de l’air microbien dans leurs services de travail: un hôpital à Bangkok. Oman Med j . (2014) 29: 346–50. doi: 10.5001 / OMJ.2014.92
43. Chughtai, AA, Stelzer-Braid, S, Rawlinson, W, Pontivivo, G, Wang, Q, Pan, Y, et al. Contamination par les virus respiratoires sur la surface extérieure des masques médicaux utilisés par les travailleurs de la santé à l’hôpital. BMC Infecte Dis . (2019) 19: 491. doi: 10.1186 / s12879-019-4109-x
44. Monalisa, D, Aruna, CN, Padma, KB, Manjunath, K, Hemavathy, E et Varsha, D. Contamination microbienne des masques buccaux utilisés par les étudiants post-diplômés dans une institution dentaire privée: une étude in vivue . J Dent Med Sci . (2017) 16: 61–7. doi: 10.9790 / 0853-1605046167
45. Kisielinski, K et Wojtasik, B. Adéposition à la coloration au sel de sodium du bengale rose pour la visualisation de la contamination du masque facial par les organismes vivants. Vise . (2022) 9: 218–31. doi: 10.3934 / Environsci.2022015
46. Liu, Z, Chang, Y, Chu, W, Yan, M, Mao, Y, Zhu, Z, et al. Masques chirurgicaux comme source de contamination bactérienne pendant les procédures opératoires. J orthopédique trad . (2018) 14: 57–62. doi: 10.1016 / j.jot.2018.06.002
47. Direction générale pour la santé et les consommateurs (Commission européenne) maintenant connue sous le nom de l’évaluation des risques plus pertinente pour la gestion des risques, Publications Office of the European Union, Lu. (2013). Disponible sur: https://data.europa.eu/doi/10.2772/34776 (consulté le 30 avril 2023).
48. Lane, DJ. Séquençage d’ARNr 16S / 23 dans: E Stackenbrandt et M Goodfellow, éditeurs. Techniques d’acide nucléique dans la systématique bactérienne . Chichester, Angleterre: John Wiley & Sons (1991). 115–76.
49. Turner, S, Pryer, KM, Miao, VP et Palmer, JD. Étude des relations phylogénétiques profondes entre les cyanobactéries et les plastes par une petite analyse de séquence d’ARNr de sous-unité. J Eukaryot Microbiol . (1999) 46: 327–38. doi: 10.1111 / j.1550-7408.1999.tb04612.x
50. Monciardini, P, Sosio, M, Cavaletti, L, Chiocchini, C et Donadio, S. Nouvelles amorces de PCR pour l’amplification sélective de l’ADNr 16S de différents groupes d’actinomycètes1. FEMS Microbiol Ecol . (2002) 42: 419–29. doi: 10.1111 / j.1574-6941.2002.tb01031.x
51. Huang, X, Lin, J et Demner-Fushman, D. Évaluation du PICO en tant que représentation des connaissances pour les questions cliniques. Amia annu sympaths proc . (2006) 2006: 359–63.
52. Calc | LibreOffice – Suite de bureau gratuite – basée sur OpenOffice – compatible avec Microsoft. (2023). Disponible sur: https://www.libreoffice.org/discover/calc (consulté le 17 mars 2023).
53. Feenstra, RPG et Tseng, Scg. Qu’est-ce qui est réellement taché par Rose Bengal? Arch Ophthalmol . (1992) 110: 984–93. doi: 10.1001 / archopht.1992.01080190090035
55. MANVAL, nous. Tachement des bactéries et des levures avec des colorants acides. Teinter Technol . (1941) 16: 13–9. doi: 10.3109 / 10520294109106189
56. Saha, DC. Une méthode de coloration rapide pour la détection des champignons endophytes dans le gazon et les graminées fourragères. Phytopathologie . (1988) 78: 237. doi: 10.1094 / phyto-78-237
57. Wojtasik, B, Zbawicka, M, Grabarczyk, L et Juzwa, W. Flow Cytométrique Approche pour évaluer l’impact de la libération de composés en béton hydro-technique au microbiome d’eau douce. Environ monit évaluer . (2021) 193: 698. doi: 10.1007 / s10661-021-09481-5
61. Kämpfer, P, Albrecht, A, Buczolits, S et Busse, HJ. Psychrobacter faecalis sp. nov., une nouvelle espèce d’une bioaérosol provenant de fèces de pigeons. Microbiol système Syst . (2002) 25: 31–6. doi: 10.1078 / 0723-2020-00099
62. Deschaght, P, Janssens, M, Vaneechouttte, M et Wauters, G. Psychrobacter Les isolats d’origine humaine, autres que Psychrobacter phénylpyruvicus , sont principalement psychrobacter faecalis et psychrobacter pulmonis , avec une description modifiée de P. faecalis . Int J Syst Evol Microbiol . (2012) 62: 671–4. doi: 10.1099 / ijs.0.032631-0
63. Park, Am, Khadka, S, Sato, F, Omura, S, Fujita, M, Hashwaki, K, et al. Isolement bactérien et fongique des masques faciaux sous la pandémie Covid-19. scientifiques Scientifiques (2022) 12: 1 Doi: 10.1038 / s41598-022-15409-x
64. Sachdev, R, Garg, K, Singh, G et Mehrotra, V. La sauvegarde est-elle compromise? Masque en bouche chirurgical hébergeant des micro-organismes dangereux dans la pratique dentaire. J Family Med Prim Care . (2020) 9: 759–63. doi: 10.4103 / jfmpc.jfmpc_1039_19
65. Gund, MP, Boros, G, Hannig, M, Thieme-Ruffing, S, Gärtner, B, Rohrer, Tr, et al. Contamination bactérienne de la peau du front et du masque chirurgical dans le traitement dentaire produisant des aérosols. J Oral Microbiol . (2021) 13: 1978731. doi: 10.1080 / 20002297.2021.1978731
66. Gund, MP, Naim, J, Hannig, M, Halfmann, A, Gärtner, B, Boros, G, et al. Le CHX et un bouclier facial ne peuvent pas empêcher la contamination des masques chirurgicaux. Front Med (Lausanne) . (2022) 9: 896308. doi: 10.3389 / fmed.2022.896308
67. Checkchi, V, Montevecchi, M, Valeriani, L et Checchi, L. Variation Bioburden des respirateurs de la pièce du visage filtrant au fil du temps: une étude préliminaire. Matériaux . (2022) 15: 8790. doi: 10.3390 / ma15248790
68. Yousefimashouf, M, Yousefimashouf, R, Alikhani, MS, Hashemi, H, Karami, P, Rahimi, Z, et al. Évaluation de la contamination bactérienne des masques faciaux portés par le personnel dans un centre de patients hospitalisés covid 19: une étude transversale. Nouveaux microbes Nouvelles infects . (2023) 52: 101090. doi: 10.1016 / j.nmni.2023.101090
69. Nightingale, M, Mody, M, Rickard, AH et Cassone, M. Contamination bactérienne sur les masques de visage utilisés chez les soins de santé des maisons de soins infirmiers. Antimicrob Steward Healthc Epidemiol . (2023) 3: E54. doi: 10.1017 / ash.2023.130
70. Keri, VC, Kumar, A, Singh, G, Mandal, A, Ali, H, Ranjan, P, et al. Étude pilote sur le fardeau de la contamination fongique dans les masques faciaux: Besoin d’une meilleure hygiène du masque à l’ère Covid-19. Infez Med . (2021) 29: 557–61. doi: 10.53854 / liim-2904-8
71. Merad, Y, Belmokhtar, Z, Hadjazi, O, Belkacemi, M, Matmour, D, Merad, Z, et al. Contamination fongique des masques médicaux chez les travailleurs médico-légaux de l’ère Covid19. Nouveaux microbes Nouvelles infects . (2023) 53: 101134. doi: 10.1016 / j.nmni.2023.101134
73. Rengasamy, S, Miller, A, Eimer, BC et Shaffer, Re. Performance de filtration des masques chirurgicaux approuvés par la FDA. J int Soc Respir Prot . (2009) 26: 54–70.
74. Fernández-Arribas, J, Moreno, T, Bartrolí, R et Eljarrat, E. Covid-19 Masques face: une nouvelle source d’exposition humaine et environnementale aux esters organophosphatés. Environ Int . (2021) 154: 106654. doi: 10.1016 / j.envint.2021.106654
75. Ostroski, P, Masiiuk, H, Kulig, P, Skoryk, A, Wcisłek, A, Jursa-Kulesza, J, et al. Les masques médicaux n’affectent pas l’équilibre acide-base, mais pourraient faciliter la transmission de Staphylococcus aureus en milieu hospitalier pendant la pandémie Covid-19. Int J Environ Res Santé publique (2023) 20: 2 Doi: 10.3390 / ijerph2
76. Sakr, A, Brégeon, F, Mège, JL, Rolain, JM et Blin, O. Staphylococcus aureus Colonisation nasale: une mise à jour sur les mécanismes, l’épidémiologie, les facteurs de risque et les infections ultérieures. Microbiol avant . (2018) 9: 2419. doi: 10.3389 / fmicb.2018.02419
77. Dietert, RR et Dietert, JM. Le superorganisme humain: utiliser des microbes pour la liberté par rapport à la peur. Appl Microbiol . (2023) 3: 883–905. doi: 10.3390 / Applmicrobiol3030061
79. Akhtar Danesh, L, Siedidi Nejad, Z, Sarmadian, H, Fooladvand, S, Van Belkum, A et Ghaznavi-Rad, E. Élimination de Staphylococcus aureus Le transport nasal chez les patients en soins intensifs abaisse les taux d’infection. Eur J Clin Microbiol Infect Dis . (2020) 39: 333–8. doi: 10.1007 / s10096-019-03729-2
80. Han, C, Shi, J, Chen, Y et Zhang, Z. Augmentation de l’évasion de l’acné causée par le port de masque de longue date pendant la pandémie Covid-19 parmi la population générale. Dermatol ther . (2020) 33: E13704. doi: 10.1111 / dth.13704
81. Bortoluzzi, P, Boneschi, V et Veraldi, S. «Masque» Tinea: une infection croissante pendant la pandémie Covid-19. Mycopathologia . (2022) 187: 141–2. doi: 10.1007 / s11046-021-00612-7
82. Silkiss, RZ, PAAP, MK et Ugradar, S. Incidence accrue de chalazion associée à l’usure du masque facial pendant la pandémie Covid-19. Am J Ophthalmol Case Rep . (2021) 22: 101032. doi: 10.1016 / j.ajoc.2021.101032
83. Akioud, W, Sabbata, S, Mozarie, Y et Oubaaz, A. Chalazion et le masque facial Usure pendant la pandémie Covid-19: Y a-t-il un lien? J Med Health Scients Eur (2023) 5: 17–9 Doi: 10.24018 / ejmed.2023.5.2.1
84. Molero-Senosiain, M, Tiew, S, Patel, A, Houben, I et Dhillon, N. Impact de l’usure du masque facial sur la kératite bactérienne. J Fr Ophtalmol . (2023) 46: E37–9. doi: 10.1016 / j.jfo.2022.04.028
85. Sakamoto, T, Terasaki, H, Yamashita, T, Shiihara, H, Funatsu, R et Uemura, A. Incidence accrue de l’endophtalmiste après vitrectomie par rapport à la porte du masque en face pendant la pandémie covide-19. Br J Ophthalmol . (2023) 107: 1472–7. doi: 10.1136 / bjophthalmol-2022-321357
86. Goncheva, MI, Gibson, RM, SHURTH, AC, Dikeakos, JD et Heinrichs, DE. L’ ISDA de la protéine Staphylococcus aureus augmente la réplication du SARS COV-2 en modulant la signalisation JAK-Stat. iscience . (2023) 26: 105975. doi: 10.1016 / j.isci.2023.105975
88. Schoenfelder, Smk, Lange, C, Eckart, M, Hennig, S, Kozytska, S et Ziebuhr, W. Succès à travers la diversité – comment Staphylococcus epidermidis établit comme un pathogène nosocomial. Int J Med Microbiol . (2010) 300: 380–6. doi: 10.1016 / j.ijmm.2010.04.011
90. Khorvash, F, Abdi, F, Kashani, HH, Naeini, FF et Nariman, T. Staphylococus aureus dans la pathogenèse de l’acné: une étude cas-témoins. N Am J Med Scients (2012) 4: 573–6 Doi: 10.4103 / 1947-2
91. Findley, K et Grice, Ea. Le microbiome cutané: l’accent mis sur les agents pathogènes et leur association avec les maladies de la peau. PHOST PATHOG . (2014) 10: E1004436. doi: 10.1371 / journal.ppat.1004436
92. Bjerre, Rd, Bandier, J, Skov, L, Engstrand, L et Johansen, Jd. Le rôle du microbiome cutané dans la dermatite atopique: une revue systématique. Br J Dermatol . (2017) 177: 1272–8. doi: 10.1111 / bjd.15390
93. Foo, CCI, Goon, ATJ, Leow, Y et Goh, C. Réactions cutanées indésirables à l’équipement de protection personnelle contre le syndrome respiratoire aigu sévère – une étude descriptive à Singapour. Contactez Derm . (2006) 55: 291–4. doi: 10.1111 / j.1600-0536.2006.00953.x
94. Rosner, E. Effets indésirables de l’utilisation prolongée du masque chez les professionnels de la santé pendant Covid-19. J Infect Dis Epidemiol . (2020) 6: 130. doi: 10.23937 / 2474-3658 / 1510130
95. Techasatian, L, Lebing, S, Uppala, R, Thaowandee, W, Chaiyarit, J, Supakunpinyo, C, et al. Les effets du masque facial sur la peau en dessous: une étude prospective pendant la pandémie Covid-19. J Prim Care Community Health . (2020) 11: 2150132720966167. doi: 10.1177 / 2150132720966167
96. Abduljabbar, M, Kalthoum, DE, Bakarman, M, Wahby Salem, I, Alsulaimani, Z, Alharbi, W, et al. La corrélation entre le port de masques du visage et les dommages cutanés chez les adultes pendant la pandémie Covid-19: une étude transversale à Djeddah, Arabie saoudite. Cureus . (2022) 14: E31521. doi: 10.7759 / Cureus.31521
98. Bakhsh, RA, Saddeeg, Sy, Basaqr, KM, Alshammrani, BM et Zimmo, Bs. Prévalence et facteurs associés de l’acné induite par le masque (Maskne) dans la population générale de la Jeddah pendant la pandémie Covid-19. Cureus . (2022) 14: E26394. doi: 10.7759 / Cureus.26394
99. Dani, A, Eseonu, A et Bibee, K. Facteurs de risque pour le développement de l’acné chez les travailleurs de la santé pendant la pandémie Covid-19. Arch Dermatol Res . (2023) 315: 1067–70. doi: 10.1007 / s00403-022-02434-Z
100. Falodun, O, Medugu, N, Sabir, L, Jibril, I, Oyakhire, N et Adekeye, A. Une étude épidémiologique sur les masques faciaux et l’acné dans une population nigériane. Plos un . (2022) 17: E0268224. doi: 10.1371 / journal.pone.0268224
102. Tallent, Sandra M., Knolhoff, Ann, Rhodehamel, E. Jeffery, Harmon, Stanley M. et Bennett, Reginald W., Bacteriological Analytical Manual (BAM); Chapitre 14: Bacillus cereus ., Fda. (1996). Disponible sur: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-14-bacillus-cenreus (consulté le 15 octobre 2023).
103. Roberge, R, Benson, S et Kim, JH. Charge thermique des respirateurs de la pièce de face filtrante N95. Ann occupe Hyg . (2012) 56: 808–14. doi: 10.1093 / annhyg / mes001
104. Roberge, RJ, Kim, JH et Benson, Sm. Absence de changements conséquents dans les réponses physiologiques, thermiques et subjectives du port d’un masque chirurgical. Respir Physiol Neurobiol . (2012) 181: 29–35. doi: 10.1016 / j.resp.2012.01.010
105. Kim, JH, Benson, SM et Roberge, RJ. Réponses pulmonaires et de la fréquence cardiaque au port de respirateurs N95 Filtrage Facepie. Am J Contrôle infecté . (2013) 41: 24–7. doi: 10.1016 / j.ajic.2012.02.037
106. Scarano, A, Inchingolo, F et Lorusso, F. Température et inconfort de la peau faciale lorsque vous portez des masques de visage protecteurs: évaluation de l’imagerie infrarouge thermique et mains en mouvement du masque. Int J Environ Res Santé publique . (2020) 17: 4624. doi: 10.3390 / ijerph17134624
107. Park, Sr, Han, J, Yeon, YM, Kang, NY et Kim, E. Effet du masque facial sur les caractéristiques cutanées changent pendant la pandémie Covid-19. Skin Res Technol . (2021) 27: 554–9. doi: 10.1111 / srt.12983
108. Lee, YH, Kim, H, Heo, DW, Ahn, IS, et Park, HK. Microbiome oral de la surface intérieure des masques faciaux et de la salive entière pendant la pandémie Covid-19. Santé orale avant . (2023) 4: 1178020. doi: 10.3389 / Froh.2023.1178020
109. Szunerits, S, Dӧrfler, H, Pagneux, Q, Daniel, J, Wadekar, S, Woitrain, E, et al. Le condensat de la respiration expirée en tant que bioanalyte: des considérations de collecte à la détection des biomarqueurs. Anal Bioanal Chem . (2023) 415: 27–34. doi: 10.1007 / s00216-022-04433-5
110. Xiang, G, Xu, K, Jian, Y, He, L, Shen, Z, Li, M, et al. Un masque prolongé portant une caractérisation microbienne nasale changée des jeunes adultes pendant la pandémie Covid-19 à Shanghai, en Chine. Immunol avant . (2023) 14: 1266941. doi: 10.3389 / fimmu.2023.1266941
111. Viola, IM, Peterson, B, Pisetta, G, Pavar, G, Akhtar, H, Menoloascina, F, et al. Revêtements de face, dispersion des aérosols et atténuation du risque de transmission du virus, IEEE Open J Eng. Med Biol . (2021) 2: 26–35. doi: 10.1109 / ojemb.2021.3053215
112. Jia, Z, Ai, Z, Cao, S et Bekö, G. Efficacité de l’équipement de protection respiratoire sur le contrôle de la source des polluants expirés. J Build Eng . (2024) 86: 108742. doi: 10.1016 / j.jobe.2024.108742
113. Barari, K, Si, X et Xi, J. Impacts de l’usure des masques et des fuites sur les flux respiratoires cycliques et la thermorégulation faciale. Fluides . (2024) 9: 9. doi: 10.3390 / fluids9010009
115. Shah, Y, Kurelek, JW, Peterson, SD et Yarusevych, S. Investigation expérimentale de la dispersion et de l’accumulation d’aérosols intérieurs dans le contexte de Covid-19: Effets des masques et de la ventilation. Fluides physiques . (2021) 33: 073315. doi: 10.1063 / 5.0057100
116. Datta, R. Utilisation de masques de facettes chirurgicales dans le théâtre de l’opération: efficace ou habitude? Med J Forces armées en Inde . (2010) 66: 163–5. doi: 10.1016 / s0377-1237 (10) 80133-9
117. Barbosa, MH, et Graziano, Ku. Influence du temps de port sur l’efficacité des masques chirurgicaux jetables comme barrière microbienne. Braz J Microbiol . (2006) 37: 216–7. doi: 10.1590 / s1517-83822006000300003
118. Kelkar, États-Unis, Gogate, B, Kurpad, S, Gogate, P et Deshpande, M. Quelle est l’efficacité des masques du visage dans le théâtre de l’opération? Une analyse du calendrier et des recommandations. INT J Contrôle d’infection . (2013) 9: 1–6. doi: 10.3396 / ijic.v9i1.003.13
119. Tcharkhtchi, A, Abbasnezhad, N, Zarbini Seydani, M, Zirak, N, Farzaneh, S et Shirinbayan, M. Un aperçu de l’efficacité de filtration à travers les masques: mécanismes de la pénétration des aérosols. Bioact Mater . (2021) 6: 106–22. doi: 10.1016 / j.bioactmat.2020.08.002
120. McCullough, NV, Brosseau, LM et Vesley, D. Collection de trois aérosols bactériens par respirateur et masque chirurgical filtres dans des conditions de flux et d’humidité relative. Ann occupe Hyg . (1997) 41: 677–90. doi: 10.1016 / s0003-4878 (97) 00022-7
121. Hadayer, A, Zahavi, A, Livny, E, Gal-Or, O, Gershoni, A, Mimouni, K, et al. Les patients portant des masques faciaux lors des injections intravitréennes peuvent être à un risque plus élevé d’endophtalmiste. Rétine . (2020) 40: 1651–6. doi: 10.1097 / iae.000000000000002919
122. Huber, C. Masques, fausse sécurité et dangers réels, partie 4: Mécanismes proposés par lesquels les masques augmentent le risque de Covid-19. Docteur principal Med J. (2020) 1: 1–9. doi: 10.6084 / m9.figshare.14021057
123. Borovoy, B, Huber, C et Crisler, M. Masks, False Safety and Real Dangers, Partie 2: Défis microbiens des masques. PDMJ . (2020) 1: 1–19. Disponible sur: https://pdmj.org/masks2/mask_risks_part2.pdf
124. Wyszyńska, M, Czelakowska, A, Rosak, P, Białożyt-Bujak, E, Gruca, O, Rosak-Szyrocka, J, et al. Changements dans la surface des muqueuses de la cavité buccale sous l’influence du port de masques de visage protectrice – analyse de concentration en oxyde nitrique – rapport préliminaire. Revêtements . (2022) 12: 1164. doi: 10.3390 / revêtements12081164
125. ICRP: Modèle des voies respiratoires humains pour la protection radiologique. Un rapport d’un groupe de travail de la Commission internationale sur la protection radiologique. Ann icrp . (1994) 24: 1–482.
126. Everard, ML, Hardy, JG et Milner, AD. Comparaison du dépôt d’aérosols nébulisé dans les poumons des adultes en bonne santé après l’inhalation orale et nasale. Thorax . (1993) 48: 1045–6. doi: 10.1136 / thx.48.10.1045
127. Cengiz, C et Can, İh. L’effet du N95 et des masques chirurgicaux sur la fonction de clairance mucociliaire et les plaintes sinonasales. Eur Arch Otorrinolaringol . (2022) 279: 759–64. doi: 10.1007 / s00405-021-06838-x
128. Sangkham, S, Faikhaw, O, Munkong, N, Sakunkoo, P, Arunlertaree, C, Chavali, M, et al. Une revue sur les microplastiques et les nanoplastiques dans l’environnement: leur occurrence, les voies d’exposition, les études toxiques et les effets potentiels sur la santé humaine. Mar Pollut Bull . (2022) 181: 113832. doi: 10.1016 / j.marpolbul.2022.113832
129. Kisielinski, K, Hockertz, S, Hirsch, O, Korupp, S, Klosterhalfen, B, Schnepf, A, et al. Le port de masques pour le visage comme source potentielle d’inhalation et d’absorption orale des toxines inanimées – une revue de cadrage. Ecotoxicol Environ Saf . (2024) 275: 115858. doi: 10.1016 / j.ecoenv.2023.115858
130. Khan, A et Jia, Z. Aperçu récent de l’absorption, de la toxicité et des cibles moléculaires des microplastiques et des nanoplastiques pertinentes pour les impacts sur la santé humaine. iscience . (2023) 26: 106061. doi: 10.1016 / j.isci.2023.106061
131. Liang, H, Ji, Y, Ge, W, Wu, J, Song, N, Yin, Z, et al. Libérez la cinétique des microplastiques à partir de masques faciaux jetables dans l’environnement aqueux. Sci Total Environ . (2022) 816: 151650. doi: 10.1016 / j.scitotenv.2021.151650
132. Ma, J, Chen, F, Xu, H, Jiang, H, Liu, J, Li, P, et al. Les masques faciaux comme source de nanoplastiques et de microplastiques dans l’environnement: quantification, caractérisation et potentiel de bioaccumulation. Environ Pollut . (2021) 288: 117748. doi: 10.1016 / j.envpol.2021.117748
133. Zhang, M, Liu, T, Zhang, L, Hua, Z, Guo, Z, Dong, J, et al. Évaluation de l’exposition microplastique dans le liquide de lavage nasal et l’influence des masques faciaux. J Hazard Mater . (2024) 480: 136069. doi: 10.1016 / j.jhazmat.2024.136069
134. Wieland, S, Balmes, A, Bender, J, Kitzinger, J, Meyer, F, Ramsperger, AF, et al. Des propriétés à la toxicité: comparaison des microplastiques à d’autres microparticules aéroportées. J Hazard Mater . (2022) 428: 128151. doi: 10.1016 / j.jhazmat.2021.128151
135. Kasloff, SB, Leung, A, Strong, JE, Funk, D et Cutts, T. Stabilité du SARS-COV-2 sur l’équipement de protection individuelle critique. Sci Rep . (2021) 11: 984. doi: 10.1038 / s41598-020-80098-3
137. Sawada, Y. Dermatite cutanée professionnelle chez les agents de santé associés à la pandémie Covid-19: une revue de la littérature. Int J Mol Sci . (2023) 24: 2989. doi: 10.3390 / ijms24032989
138. Tunçer Vural, A. Le développement d’acné vulgaris en raison de masques face pendant la pandémie, de sensibilisation au risque et d’attitudes d’un groupe d’étudiants universitaires. J Cosmet Dermatol . (2022) 21: 5306–13. doi: 10.1111 / jocd.15120
139. Liu, Y, Zhao, H, Chen, H, Li, X, Ran, C, Sun, H, et al. Le port du masque affecte-t-il la santé de la peau? Une étude de métabolomique cutanée non ciblée. Environ Int . (2023) 178: 108073. doi: 10.1016 / j.envint.2023.108073
140. Brooks, JK, Sultan, AS, et Jabra-Rizk, MA. L’usure prolongée du masque facial est une préoccupation pour le développement du microbiome dysbiotique. Respirat Med Res . (2022) 81: 100877. doi: 10.1016 / j.resmer.2021.100877
141. Koshevarova, VA, Westenhaver, ZK, Schmitz-Brown, M, McKinnon, BJ, Merkley, KH et Gupta, PK. Blépharoconjunctivitis et tendances des maladies oto-rhino-laryngologiques dans le contexte de la port de masques pendant la pandémie Covid-19. CLIN PRACT . (2022) 12: 619–27. doi: 10.3390 / clinpract12040065
142. Schulttheis, WG, Sharpe, JE, Zhang, Q, Patel, Sn, Kuriyan, AE, Chiang, A, et al. Effet de l’enregistrement des masques de face sur le nombre quantitatif de particules près de l’œil: implications pour les injections intravitréennes dans l’ère Covid-19. Am J Ophthalmol . (2021) 225: 166–71. doi: 10.1016 / j.ajo.2021.01.021
143 Utilisation du masque facial et effets sur la santé de la surface oculaire: une revue complète. OCUL SURF . (2023) 27: 56–66. Doi: 10.1016 / j.jtos.2022.12,006
146. Islam, Sr, Prusty, D, Maiti, S, Dutta, R, Chattopadhyay, P et Manna, Skk. Effet de l’utilisation à court terme du masque FFP2 (N95) sur le métabolome salivaire des jeunes volontaires sains: une étude pilote. Mol omics . (2023) 19: 383–94. doi: 10.1039 / d2mo00232a
147. Arora, U, Priyadarsh, M, Katiyar, V, Sonoja, M, Garg, P, Gupta, I, et al. Facteurs de risque de mucormycose associée à la maladie du coronavirus. J Infecter (2022) 84: 383–90. Doi: 10.1016 / j.2021.12.03
148. Kisielinski, K, Steigleder-Schweiger, C, Wagner, S, Korupp, S, Hockertz, S et Hirsch, O. Risques et avantages des masques faciaux chez les enfants. Préimprimés . (2024) 1–51. doi: 10.20944 / prerints202409.1508.v1
150. Matuschek, C, Moll, F, Fangerau, H, Fian, JC, Sönger, K, de Griennven, M, et al. L’histoire et la valeur ou les masques faciaux. Eur J Med Res . (2020) 25:23. doi: 10 1186 / S40001-020-00423-4
151. Lee, SA, Grinshpun, SA et Reponen, T. Performance respiratoire offerte par les respirateurs N95 et les masques chirurgicaux: évaluation du sujet humain avec aérosol NaCl représentant la gamme de taille des particules bactériennes et virales. Ann occupe Hyg . (2008) 52: 177–85. doi: 10.1093 / Annhyg / Men005
152. Ntlailane, MGL et Wichmann, J. Efficacité des respirateurs de N95 pour le contrôle de l’exposition aux nanoparticules (2000-2016): une revue systématique et une méta-analyse. J Nanopart Res . (2019) 21: 170. doi: 10.1007 / s11051-019-4596-0
153. Samaranayake, LP, Fakhruddin, KS, ONG, HC, Chang, JWW et Panduwawala, C. L’efficacité et l’efficacité de l’équipement de protection respiratoire (RPE) en dentisterie et autres établissements de soins de santé: un examen systématique. Acta Odontol Scand . (2020) 78: 626–39. doi: 10.1080 / 00016357.2020.1810769
154. Willeke, K, Qian, Y, Donnelly, J, Grinshpun, S et Ulevicius, V. Pénétration des micro-organismes aéroportés à travers un masque chirurgical et un respirateur de poussière / brume. Am Ind Hyg Assoc j . (1996) 57: 348–55. doi: 10.1080 / 15428119691014882
156. Qian, Y, Willeke, K, Grinshpun, SA, Donnelly, J et Coffey, CC. Performance des respirateurs N95: efficacité de filtration pour les particules microbiennes et inertes aériennes. Am Ind Hyg Assoc j . (1998) 59: 128–32. doi: 10.1080 / 15428119891010389
157. Loeb, M, Dafoe, N, Mahony, J, John, M, Sarabia, A, Glavin, V, et al. Masque chirurgical vs respirateur N95 pour prévenir la grippe chez les travailleurs de la santé: un essai randomisé. Jama . (2009) 302: 1865–71. doi: 10.1001 / jama.2009.1466
158. Smith, JD, MacDougall, CC, Johnstone, J, Copes, RA, Schwartz, B et Garber, Ge. Efficacité des respirateurs N95 par rapport aux masques chirurgicaux dans la protection des travailleurs de la santé contre les infections respiratoires aiguës: une revue systématique et une méta-analyse. CMAJ . (2016) 188: 567–74. doi: 10.1503 / CMAJ.150835
159. Liu, I, Prasad, V et Darrow, J. Preuve du masquage du visage communautaire pour limiter la propagation de SAR-COV-2: une revue critique. Matrice de santé J Law Med . (2023) 33: 1. Disponible sur: https://scholarlycommons.law.case.edu/healthmatrix/vol33/iss1/1/
160. Vincent, M, et Edwards, P. masques de visage chirurgical jetable pour prévenir l’infection chirurgicale en chirurgie propre. COCHRANE DATABASE SYST REV . (2016) 2016: CD002929. doi: 10.1002 / 14651858.cd002929.pub3
161. Burdick, HN et Maibach, H. Pertinence clinique des masques dans la salle d’opération? Une revue systématique. Clin Infect Pract . (2021) 12: 100087. doi: 10.1016 / j.clinpr.2021.100087
162. Carbon, CC. Le port de masques pour le visage confond fortement les homologues dans la lecture des émotions. Front Psychol . (2020) 11: 566886. doi: 10.3389 / fpsyg.2020.566886
164. OMS, World Medical Association (WMA): Déclaration d’Helsinki. Principes éthiques pour la recherche médicale impliquant des sujets humains. Bull World Health Organ . (2001) 79: 373–4.
165. Elgersma, IH, Fretheim, A, Elstrøm, P et Aavitsland, P. Association entre l’utilisation du masque facial et le risque d’infection SAR-COV-2: étude transversale. Infection de l’épidémiol . (2023) 151: E194. doi: 10.1017 / s0950268823001826
166. Boretti, A. Efficacité des mandats de masquage du visage généralisés, de la recherche sur les services de santé et de la gestion. Épidémiologie . (2021) 8: 23333928211058023. doi: 10.1177 / 23333928211058023
167. Galanis, P, Vraka, I, Fragkou, D, Bilali, A et Kaitelidou, D. Impact de l’utilisation des équipements de protection personnelle sur la santé physique des travailleurs de la santé pendant la pandémie de Covid-19: une revue systématique et une méta-analyse des travailleurs . Am J Contrôle infecté . (2021) 49: 1305–15. doi: 10.1016 / j.ajic.2021.04.084
168. Unoki, T, Sakuramoto, H, Sato, R, Ouchi, A, Kuribara, T, Furumaya, T, et al. Effets négatifs de l’équipement de protection personnelle parmi les professionnels de la santé des unités de soins intensifs pendant la pandémie Covid-19: une revue de portée. Sage Open Nurse . (2021) 7: 23779608211026164. doi: 10.1177 / 23779608211026164
169. Dirol, H, Alkan, E, Sindel, M, Ozdemir, T et Erbas, D. Les effets physiologiques et inquiétants des masques de visage chirurgical à l’ère Covid-19. BLL . (2021) 122: 821–5. doi: 10.4149 / bll_2021_131
170. Gaikwad, RP, Banodkar, AB et Nandgaonkar, vice-président. Conséquences respiratoires du masque N95 pendant la pandémie Covid-19 – une étude observationnelle. Int J Health Sci Res . (2021) 11: 55–61. doi: 10.52403 / ijhsr.20210407
171. Walach, H, Traindl, H, Prentice, J, Weikl, R, Diemer, A, Kappes, A, et al. Le dioxyde de carbone s’élève au-delà des niveaux de sécurité acceptables chez les enfants sous couverture du nez et de la bouche: résultats d’une étude de mesure expérimentale chez les enfants en bonne santé. Environ Res . (2022) 212: 113564. doi: 10.1016 / j.envre.2022.113564
172. Acuti Martellucci, C, Flacco, ME, Martellucci, M, Violante, FS et Manzoli, L. Concentration inhalée du CO 2 tout en portant des masques faciaux: une étude pilote utilisant la capnographie. Environ Health Insights . (2022) 16: 11786302221123573. doi: 10.1177 / 11786302221123573
173. Ahmad, MDF, Wahab, S, Ali Ahmad, F, Intakhab Alam, M, Ather, H, Siddiqua, A, et al. Une nouvelle approche en perspective pour explorer les avantages et les inconvénients du masque facial dans la prévention de la propagation du SRAS-COV-2 et d’autres agents pathogènes. Pharm saoudien j . (2021) 29: 121–33. doi: 10.1016 / j.jsps.2020.12.014
174. Shobako, N. Leçons des politiques de santé pour les enfants pendant la pandémie au Japon. Santé publique avant . (2022) 10: 1015955. doi: 10.3389 / fpubh.2022.1015955
175. Kampf, G. Effet du masquage du visage sur la transmission de SARS-CoV-2 dans: N Rezaei, éditeur. Les conséquences Covid-19: Volume II: leçons apprises . Suisse, Cham: Springer Nature (2024). 175–99. doi: 10.1007 / 978-3-031-61943-4_12
176. Beauchamp, JD, et Mayhew, ca. Revisiter la justification du masquage obligatoire. J souffle res . (2023) 17: 042001. doi: 10.1088 / 1752-7163 / ACDF12
177. Sandlund, J, Duriseti, R, Ladhani, Sn, Stuart, K, Noble, J et Beth Høeg, T. Masques face et protection contre le Covid-19 et d’autres infections respiratoires virales: évaluation des avantages et préjudices chez les enfants. Paediatr respir Rev. (2024). doi: 10.1016 / j.prrv.2024.08.003
178. Mastropasqua, L, Lanzini, M, Brescia, L, D’Aloisio, R, Nubile, M, Ciancaglini, M, et al. Modifications de surface oculaire liées au masque en face pendant la pandémie Covid-19: une microscopie confocale clinique, in vivo et une étude immuno-cytologie. Trad vis sci technol . (2021) 10:22. doi: 10.1167 / tvst.10.3.22
179. D’souza, S, Vaidya, T, Nair, AP, Shetty, R, Kumar, Nr, Bisht, A, et al. Altération de l’état de santé de la surface oculaire et profil immunitaire du film lacrymal en raison d’une usure de masque quotidienne prolongée chez les travailleurs de la santé. Biomédicines . (2022) 10: 1160. doi: 10.3390 / biomédicines10051160
180. Jin, S, Wetzel, D et Schirmer, M. Déchiffrer les mécanismes et implications de la translocation bactérienne en santé humaine et maladie. Curr Opin Microbiol . (2022) 67: 102147. doi: 10.1016 / j.mib.2022.102147
181. Asadi, S, Cappa, CD, Barreda, S, Wexler, AS, Bouvier, NM et RistenPart, Wd. Efficacité des masques et des revêtements de visage dans le contrôle des émissions de particules aérosols extérieures des activités expiratoires. Sci Rep . (2020) 10: 15665. doi: 10.1038 / s41598-020-72798-7
182. Bagchi, S, Basu, S, Chaudhuri, S et Saha, A. Pénétration et atomisation secondaire des gouttelettes touchées sur les masques humides. Phys Rev Fluids . (2021) 6: 110510. doi: 10.1103 / PhysRevfluids.6.110510
183. Rebmann, T, Carrico, R et Wang, J. Effets physiologiques et autres et conformité à l’utilisation des respirateurs à long terme chez les infirmières de l’unité de soins intensifs médicaux. Am J Contrôle infecté . (2013) 41: 1218–23. doi: 10.1016 / j.ajic.2013.02.017
184. Matusiak, ł, Szepietowska, M, Krajewski, P, Białynicki-Birula, R, et Szepietowski, JC. Inconvénients dus à l’utilisation de masques faciaux pendant la pandémie Covid-19: une étude de 876 jeunes. Dermatol ther . (2020) 33: E13567. doi: 10.1111 / dth.13567
185. Naylor, G, Burke, LA et Holman, Ja. Le verrouillage de Covid-19 affecte le handicap auditif et le handicap de diverses manières: une étude d’enquête en ligne rapide. Oreille entendre . (2020) 41: 1442–9. doi: 10.1097 / aud.0000000000000948
186. Thomas, F, Allen, C, Butts, W, Rhoades, C, Brandon, C et Handrahan, DL. Le port d’un masque chirurgical ou de N95-Ruirator altait-il la communication radio? Air Med J. (2011) 30: 97–102. doi: 10.1016 / j.amj.2010.12.007
187. Heider, CA, Álvarez, ML, Fuentes-López, E, González, CA, León, Ni, Verástegui, DC, et al. Prévalence des troubles de la voix chez les travailleurs de la santé dans l’ère universelle du masquage Covid-19. Laryngoscope . (2020) 131: E1227–33. Doi: 10.1002 / Lary.
188. Zarei, N, Negarandeh, R et Neshat, H. Défis de communication causés par le port de masques et stratégies utilisés par les infirmières pédiatriques pendant la pandémie Covid-19: une étude qualitative. J Pediatr Nurs . (2024) 77: E54–61. doi: 10.1016 / j.ppedn.2024.03.020
189. Sezer, H, çınar, D et Kılıç Akça, N. L’effet de l’utilisation prolongée des masques chirurgicaux pendant l’enseignement en face à face sur les paramètres cognitifs et physiologiques des étudiants en soins infirmiers: une étude transversale et descriptive. Nurse Educ Practice . (2023) 72: 103779. doi: 10.1016 / j.nepr.2023.103779
19 .. Elle, C, Haim, Have Grot, Jx, Sch et Junne, Yennament, H. PROVE SIGNRICAL FIMTIANT LOI ASTES SITS SITE Effective Effective J. (2021) 58: 2101131. Doi: 10.1183 / 13993003.01131-2021
191. Chandra, A et Høeg, TB. Manque de corrélation entre les mandats de masque scolaire et les cas de Covid-19 de Covid-19 pédiatriques dans une grande cohorte. J Infect . (2022) 85: 671–5. doi: 10.1016 / j.jinf.2022.09.019
193. Wang, MX, Gwee, Sxw, Chua, Pey et Pang, J. Efficacité des masques de visage chirurgical dans la réduction des infections respiratoires aiguës en milieu de santé: une revue systématique et une méta-analyse. Avant Med . (2020) 7: 564280. doi: 10.3389 / fmed.2020.564280
194. Fønhus, MS, Dalsbø, TK et Brurberg, KG, masques pour prévenir la transmission des maladies respiratoires, telles que Covid-19, Norwegian Institute of Public Health. (2021). Disponible sur: https://hdl.handle.net/11250/2756758
195. Aiello, AE, Perez, V, Coulborn, RM, Davis, BM, Uddin, M et Monto, AS. Facemases, hygiène des mains et grippe chez les jeunes adultes: un essai d’intervention randomisé. Plos un . (2012) 7: E29744. doi: 10.1371 / journal.pone.0029744
197. Alfelali, M, Haworth, EA, Barasheed, O, Badahdah, AM, Bokhary, H, Tashani, M, et al. Équipe de recherche du Hajj, Facemask contre les infections respiratoires virales chez les pèlerins du Hajj: un essai difficile en grappe. Plos un . (2020) 15: E0240287. doi: 10.1371 / journal.pone.0240287
199. MacIntyre, CR, Cauchemez, S, Dwyer, DE, Seale, H, Cheung, P, Browne, G, et al. Utilisation du masque facial et contrôle de la transmission du virus respiratoire dans les ménages. Émerger infecte dis . (2009) 15: 233–41. doi: 10.3201 / eid1502.081166
200. MacIntyre, CR, Seale, H, Dung, TC, Hien, NT, NGA, PT, Chughtai, AA, et al. Un essai randomisé en grappe de masques en tissu par rapport aux masques médicaux chez les travailleurs de la santé. BMJ ouvert . (2015) 5: E006577. doi: 10.1136 / bmjopen-2014-006577
201. Simmerman, JM, Suntarattiwong, P, Levy, J, Jarman, RG, Kaewchana, S, Gibbons, RV, et al. Résultats d’un essai contrôlé randomisé au ménage de lavage des mains et de masques faciaux pour réduire la transmission de la grippe à Bangkok, en Thaïlande. Grippe d’autres virus respir . (2011) 5: 256–67. doi: 10.1111 / j.1750-2659.2011.00205.x
202. Cowling, BJ, Fung, Rop, Cheng, Cky, Fang, VJ, Chan, KH, Seto, WH, et al. Résultats préliminaires d’un essai randomisé d’interventions non pharmaceutiques pour empêcher la transmission de la grippe dans les ménages. Plos un . (2008) 3: E2101. doi: 10.1371 / journal.pone.0002101
203. Cowling, BJ, Chan, KH, Fang, VJ, Cheng, CKY, Fung, Rop, Wai, W, et al. Facemases et hygiène des mains pour prévenir la transmission de la grippe dans les ménages: un essai randomisé en grappe. Ann Intern Med . (2009) 151: 437–46. doi: 10.7326 / 0003-4819-151-7-200910060-00142
204. Suess, T, Remschmidt, C, Schink, SB, Schweiger, B, Nitsche, A, Schroeder, K, et al. Le rôle des masques faciales et de l’hygiène des mains dans la prévention de la transmission de la grippe dans les ménages: résultats d’un essai randomisé en grappe; Berlin, Allemagne, 2009-2011. BMC Infecte Dis . (2012) 12:26. doi: 10.1186 / 1471-2334-12-26
205. Larson, El, Ferng, Y, Wong-McLoughlin, J, Wang, S, Haber, M et Morse, SS. Impact des interventions non pharmaceutiques sur les uris et la grippe dans les ménages urbains bondés. Rep . (2010) 125: 178–91. doi: 10.1177 / 003335491012500206
206. Jacobs, JL, Ohde, S, Takahashi, O, Tokuda, Y, Omata, F et Fukui, T. procès. Am J Contrôle infecté . (2009) 37: 417–9. doi: 10.1016 / j.ajic.2008.11.002
207. Bundgaard, H, Bundgaard, JS, Raaschou-Pedersen, Det, von Buchwald, C, Todsen, T, Norwegian, JB, et al. Efficacité de l’ajout d’une recommandation de masque à d’autres mesures de santé publique pour prévenir l’infection SAR-COV-2 chez les porteurs de masques danois. Ann interne avec . (2020). doi: 10.7326 / m20-6817
208. Juutinen, A, Sarvikivi, E, Laukkanen-Nevala, P, et Helve, O. Les recommandations de masques face dans les écoles n’ont pas eu d’impact sur l’incidence de Covid-19 chez les enfants de 10 à 12 ans en Finlande – Analyse de régression de jointure. BMC Public Health . (2023) 23: 730. doi: 10.1186 / s12889-023-15624-9
209. Gómez-Ochoa, SA et Muka, T. La méta-analyse sur l’utilisation du masque en facemas en milieu communautaire pour prévenir la transmission des infections respiratoires ne montre aucun effet. Int J Infecte Dis . (2021) 103: 257–9. doi: 10.1016 / j.ijid.2020.11.139
Citation: Kisielinski K, Wojtasik B, Zalewska A, Livermore DM et Jurczak-Kurek A (2024) Le fardeau bactérien des masques de visage usés – Recherche et revue de la littérature. Devant. Santé publique . 12: 1460981. doi: 10.3389 / fpubh.2024.1460981
Reçu: 18 juillet 2024; Accepté: 30 octobre 2024; Publié: 03 décembre 2024.
Nous utilisons des cookies pour optimiser notre site web et notre service.
Fonctionnel
Toujours activé
Le stockage ou l’accès technique est strictement nécessaire dans la finalité d’intérêt légitime de permettre l’utilisation d’un service spécifique explicitement demandé par l’abonné ou l’utilisateur, ou dans le seul but d’effectuer la transmission d’une communication sur un réseau de communications électroniques.
Préférences
Le stockage ou l’accès technique est nécessaire dans la finalité d’intérêt légitime de stocker des préférences qui ne sont pas demandées par l’abonné ou l’utilisateur.
Statistiques
Le stockage ou l’accès technique qui est utilisé exclusivement à des fins statistiques.Le stockage ou l’accès technique qui est utilisé exclusivement dans des finalités statistiques anonymes. En l’absence d’une assignation à comparaître, d’une conformité volontaire de la part de votre fournisseur d’accès à internet ou d’enregistrements supplémentaires provenant d’une tierce partie, les informations stockées ou extraites à cette seule fin ne peuvent généralement pas être utilisées pour vous identifier.
Marketing
Le stockage ou l’accès technique est nécessaire pour créer des profils d’utilisateurs afin d’envoyer des publicités, ou pour suivre l’utilisateur sur un site web ou sur plusieurs sites web ayant des finalités marketing similaires.














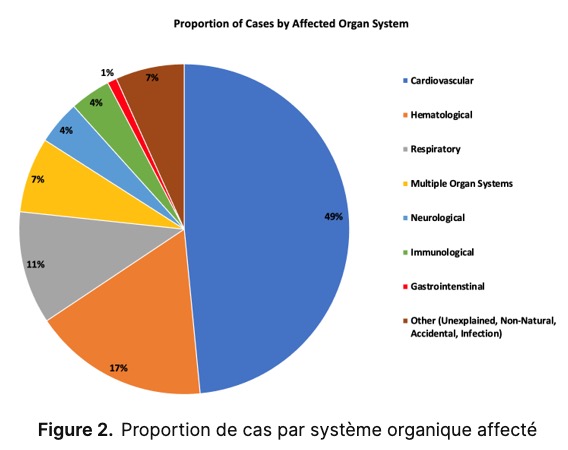



















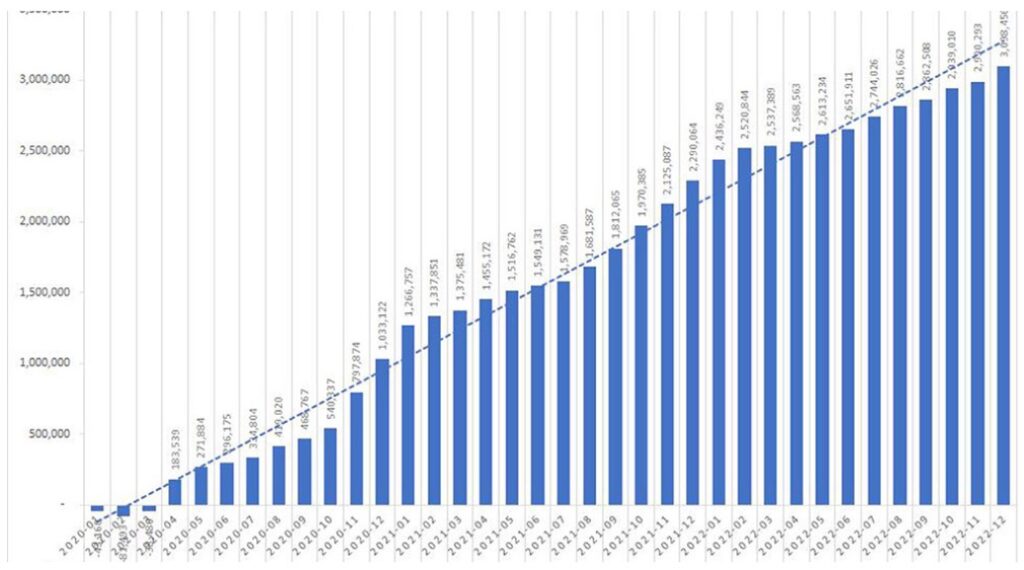
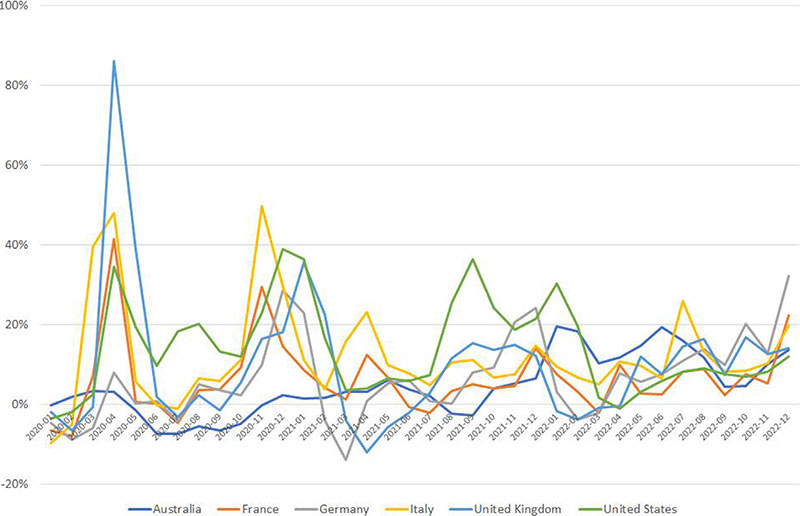






 Barbara Wojtasik2
Barbara Wojtasik2